

Summary
Early in development, primordial germ cells (PGCs) are set aside from somatic cells and acquire a unique gene expression program [1]. The mechanisms underlying germline-specific gene expression are largely unknown. Nanos expression is required during germline development [2–5] and posttranscriptionally restricted to PGCs [4, 6, 7, 8]. Here we report that the microRNA miR-430 targets the 3′ untranslated region (UTR) of nanos1 during zebrafish embryogenesis. A miR-430 target site within the nanos1 3′UTR reduces poly(A) tail length, mRNA stability and translation. Repression is disrupted in maternal-zygotic dicer mutants (MZdicer), which lack mature miRNAs [9], and restored by injection of processed miR-430. Although miR-430 represses other genes equally in germline and soma, specific regions in the nanos1 3′UTR compensate for microRNA-mediated repression in PGCs and allow germline-specific expression. We show that the 3′UTR of an additional PGC-specific gene, TDRD7, is also targeted by miR-430. These results indicate that miR-430 targets the 3′UTRs of germline genes and suggest that differential susceptibility to microRNAs contributes to tissue-specific gene expression.
Results
nanos1 3′UTR induces mRNA deadenylation
Zebrafish nanos1 mRNA is maternally supplied to the early embryo [4]. Multiple post-transcriptional mechanisms function via the nanos1 3′UTR to ensure protein expression specifically in PGCs. First, nanos1 mRNA is localized to the germ plasm at the cleavage furrows [4, 10, 11]. Second, the nanos1 3′UTR represses nanos1 translation in somatic cells. Third, nanos1 mRNA is rapidly degraded in somatic cells but stably maintained in PGCs [4]. These mechanisms lead to protein expression specifically in PGCs [4, 12]. A GFP reporter mRNA containing the nanos1 3′UTR injected at the one-cell stage is stabilized in PGCs but is translationally repressed and degraded in somatic cells [4] (Fig.1A). We took advantage of this injection assay to elucidate the molecular basis of posttranscriptional regulation of nanos1.
Figure 1. The nanos1 3′UTR induces deadenylation.

(A) GFP mRNAs with different 3′UTR fragments and control DsRed mRNA were co-injected at the one-cell stage. GFP (green) and DsRed (red) expression were analyzed at 26 hours post fertilization (hpf). The levels of GFP mRNA were examined by in situ hybridization (dark purple). Insets show the gonad region. Arrowheads indicate PGCs. (B) Schematic representation of the poly(A) tail assay for GFP reporter mRNA. See materials and methods for details. (C) Poly(A) tail profile of GFP reporter mRNAs with SV40 poly(A) signal. Time after fertilization is indicated above each lane. (A0) shows completely deadenylated fragments. Poly(A) tail length is shown on the right of each panel. (D) Poly(A) tail profile of GFP reporter mRNAs with SV40 poly(A) signal+poly(A) tail.
The poly(A) tail is a key determinant of mRNA stability and translation efficiency [13–15]. We therefore asked if the post-transcriptional repression activity of the nanos1 3′UTR in somatic cells is mediated by the control of poly(A) tail length (Fig.1B). We compared the poly(A) tail dynamics of GFP-nanos1 mRNA to two control mRNAs that contained either GFP ORF alone (GFP mRNA) or GFP ORF with the vasa 3′UTR (GFP-vasa mRNA). We used the latter reporter, because the vasa 3′UTR restricts the mRNA to PGCs but does not repress protein synthesis in somatic cells as efficiently as the nanos1 3′UTR (Fig.1A and [16] ). This suggests that the nanos1 and vasa 3′UTRs mediate posttranscriptional regulation by different mechanisms. Analysis of the poly(A) tail dynamics revealed that GFP-nanos1 mRNA was initially polyadenylated but almost completely lost its poly(A) tail at 4–6 hour post fertilization (hpf). In contrast, GFP and GFP-vasa mRNA retained a ~150 nt poly(A) tail (Fig.1C). The injected GFP reporters recapitulated the poly(A) tail dynamics of endogenous nanos1 mRNA and vasa mRNAs (Fig.S1). These results indicate that the bulk of nanos1 mRNA lost its poly(A) tail during embryogenesis due to a sequence element within its 3′UTR. Similar results were obtained when the poly(A) tail was added in vitro prior to injection (Fig.1D), suggesting that the nanos1 3′UTR induces deadenylation and might also reduce de novo polyadenylation.
Deadenylation of nanos1 mRNA is not caused by translational repression
Deadenylation could be either the cause or the result of translational repression [17]. To distinguish between these possibilities, we analyzed the poly(A) length of GFP reporter mRNAs primed with A-cap. A-cap does not interact with the translation initiation factor eIF4E and inhibits translation initiation [18]. A-capped GFP mRNA was not translated in zebrafish embryos (Fig.S2A) and had the same poly(A) profile as the m7G-capped GFP mRNA (Fig.S2B, lower panel). Conversely, A-capped GFP-nanos1 mRNA was rapidly deadenylated, similarly to m7G-capped GFP-nanos1 mRNA (Fig.S2B, upper panel). These results indicate that deadenylation promoted by the nanos1 3′UTR is not caused by the lack of translation initiation and does not require active translation.
A short cis-regulatory element controls deadenylation and translational repression
To identify the cis-regulatory element within the nanos1 3′UTR required for deadenylation, we examined the poly(A) tail dynamics and GFP expression of a series of reporters that contained deletion mutants of the nanos1 3′UTR (Fig.2A). This analysis led to the identification of a 79-nucleotide (nt) deadenylation element (Ib). A GFP reporter mRNA bearing the fragment Ib underwent deadenylation and translational repression. Conversely, deletion of the fragment Ib from the nanos1 3′UTR (ΔIb) blocked rapid deadenylation and repression of the GFP reporter in somatic cells (Fig.2A, B and not shown). Thus, the fragment Ib is necessary and sufficient to induce rapid deadenylation and repress GFP reporter expression in somatic cells.
Figure 2. Identification of the deadenylation element.
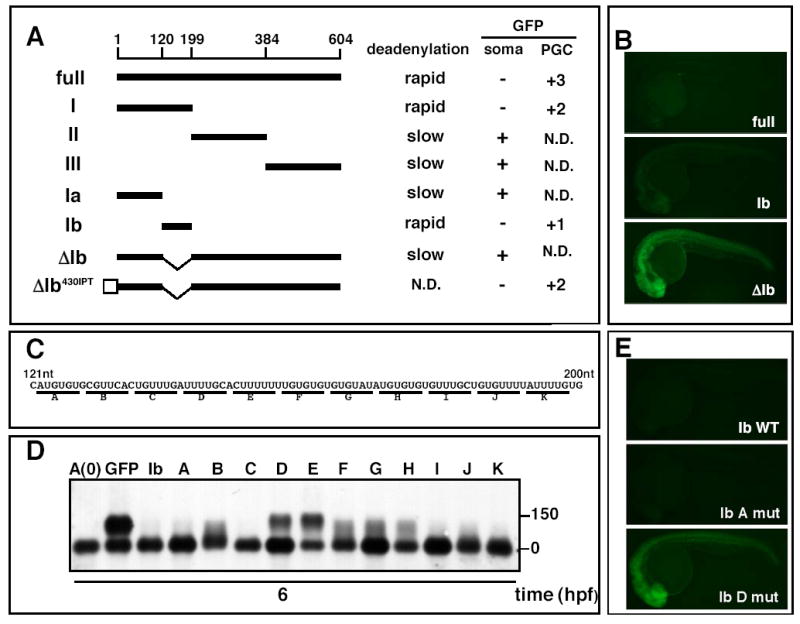
(A) Schematic representation of nanos1 3′UTR deletion mutants. Deadenylation activity (rapid or slow), GFP expression levels in somatic cells (+ or −) and PGCs (+3 represents the GFP expression of full length nanos1 3′UTR) are indicated on the right. White box indicates miR-430 site. N.D: not determined. The positions relative to the stop codon are shown above. (B) GFP fluorescence from nanos1 3′UTR deletion mutant reporters at 26 hpf. (C) Sequence of the Ib region. Base substitutions (A-K) are shown below. (D) Poly(A) profile of Ib base-substitution mutant reporters at 6 hpf. (E) GFP fluorescence of Ib mutant reporters at 26 hpf.
To identify the core sequence elements responsible for the rapid deadenylation, we used a series of mutants within region Ib (Fig.2C). Base-substitutions within regions D and E strongly inhibited deadenylation of the GFP reporter mRNA, whereas other substitutions only affected deadenylation weakly (B, F, G, and H) or had no effect (A, C, I, J and K) (Fig.2D). Concomitantly, repression of the GFP reporter was lost by substitutions D and E (Fig.2E and data not shown). These experiments indicate that the sequence elements D and E within the nanos1 3′UTR are necessary for the rapid deadenylation and repression of the GFP reporter in somatic cells.
MiR-430 induces deadenylation and translational repression in somatic cells
We hypothesized that a microRNA (miRNA) might bind to the D-E element and induce mRNA deadenylation and repression. Although their mechanism of action is not fully understood, miRNAs regulate target mRNA deadenylation [19–21] and inhibit protein synthesis by repressing translation [22–24]. We found that the sequence GCACUU in site D-E in the nanos1 3′UTR is complementary to nucleotides 2 to 7 of miR-430 (Fig.3A), the miRNA “seed” sequence important for target mRNA recognition [25–28]. MiR-430 is expressed ubiquitously during early embryogenesis and required for normal morphogenesis during gastrulation and brain development [9]. To determine if miR-430 induces target deadenylation and repression through the D-E region in the nanos1 3′UTR, we used three approaches. First, we injected the GFP-nanos1 reporter into wild type, MZdicer mutants, and MZdicer mutants injected with miR-430 duplex (MZdicer+miR-430)(Fig.3B). The RNaseIII enzyme Dicer is required for miRNA processing, and loss of maternal and zygotic dicer (MZdicer) results in loss of mature miRNAs, including miR-430 [9]. We found that repression and deadenylation of the GFP-nanos1 reporter were reduced in MZdicer mutants. Conversely, injection of miR-430 duplex into MZdicer mutants restored regulation of the nanos1 reporter mRNA (Fig.3C and D). In contrast, expression of the GFP-vasa reporter, which does neither contain a GCACUUU sequence nor promote deadenylation, is similar in wild-type and MZdicer embryos (Fig.S4). Second, we mutated two nucleotides within the predicted target site that disrupt the pairing with the miR-430 seed (GCACUU to GGUCUU; Fig. 3A). We found that these mutations lead to higher GFP expression levels in somatic cells and delayed deadenylation compared to the wild-type reporter (Fig.3E and F). Third, we inserted three copies of the BCDE element downstream of the GFP ORF (3×BCDE) (Fig.3G and H). 3×BCDE recapitulated the deadenylation and repression activity of the nanos1 3′UTR, whereas a mutated BCDE sequence (GCACUU to GGUCUU, 3×BCDEmut) did not. Furthermore, co-injection of a mutant miR-430b duplex with compensatory mutations that basepair with BCDEmut restored deadenylation and repression. These experiments indicated that the BCDE site is a bona fide miR-430 target site and induces deadenylation and repression of the GFP reporter.
Figure 3. miR-430 mediates deadenylation and repression of protein synthesis induced by the nanos1 3′UTR.

(A) Putative base pairing of miR-430b with nanos1 3′UTR BCDE site. Base-substitutions in the BCDE site to disrupt the putative miR-430b binding are shown in red. Mutated nucleotides within miR-430b to restore base pairing with the mutated nanos1 3′UTR are shown in blue. (B) Experimental set up to test the effect of miR-430 on GFP-nanos1 reporter using MZdicer mutants. (C, E, G) GFP reporter expression (green) and control DsRed expression (red) at 26 to 30 hpf in wild type (C, E, G), MZdicer (C) and MZdicer+miR-430(C). Inset shows the enlarged view of gonad region. (D, F, H) Poly(A) length of GFP-reporter mRNAs used in C, E, and G at different times after injection. The mutant reporters (E–H) (nanos1 mut) include the base substitutions shown in red(A).
MiRNAs not only cause translational repression but also degradation of target mRNAs[19, 29, 30]. We therefore asked whether miR-430 is also responsible for the clearance of the GFP-nanos1 reporter mRNA ([4] and Fig.1A). In situ hybridization showed that GFP-nanos1 mRNA accumulates in somatic cells in the absence of miR-430 regulation (Fig.S4). These experiments indicate that miR-430 accelerates GFP-nanos1 mRNA decay in somatic cells.
Cis-elements within the nanos1 3′UTR allow protein expression in PGCs in the presence of miRNAs
The differential regulation of the nanos1 3′UTR by miR-430 contrasts with previous studies of miR-430 targets. In particular, miR-430 is uniformly expressed during early embryogenesis, and previously analyzed miR-430 target mRNAs and synthetic miR-430 reporters are repressed equally in somatic cells and PGCs [9, 19]. Indeed, in the absence of additional sequences from the nanos1 3′UTR, the 3×BCDE reporter was repressed in the soma and germline (Fig.3E and 4C). Thus, miR-430 can be active both in the soma and PGCs, but the nanos1 3′UTR is more susceptible to repression in the soma. To determine the mechanistic basis for the differential repression by miR-430, we performed four experiments. First, we asked if differential susceptibility is unique to miR-430 or if other miRNAs act similarly. We replaced the miR-430 target site in the nanos1 3′UTR with a miR-204 imperfect target site (GFP-nanos1204 IPT mRNA). Injection of intermediate levels of the miR-204 duplex strongly repressed the GFP-nanos1204 IPT reporter in somatic cells but not in PGCs (Fig.4E). This experiment suggests that miRNAs have differential effects on the nanos1 3′UTR in the soma and germline. Second, we asked whether the location of the miR-430 target site within the nanos1 3′UTR affects its activity. We placed the miR-430 site at the 5′ or 3′end of the nanos1 3′UTR, 127 nt or 450 nt away from its original location (Fig.4A). The nanos1 3′UTR promoted protein synthesis in PGCs in the presence of the displaced miR-430 site. This result indicates that the local environment of the miR-430 site is not required for differential expression in the soma and germline (Fig.4F and G). Third, we determined if there are cis-elements in the nanos1 3′UTR that allow PGC expression in the presence of miR-430. We examined GFP expression of the nanos1 3′UTR deletion constructs in PGCs (Fig.2A and Fig.S3) and found that fragment Ib, which contains the miR-430 site, promoted GFP expression in PGCs. Similarly, the remainder of the nanos1 3′UTR (Δ1b) also promoted GFP expression in PGCs. Addition of the miR-430 site (ΔIb430IPT) showed that the ΔIb fragment directed protein synthesis in PGCs in the presence of miR-430 (Fig.S3). Hence, the nanos1 3′UTR contains at least two cis-regulatory regions that contribute to the differential repression in soma and germline. Fourth, we asked if the nanos1 3′UTR allows expression in PGCs by sequestering the target mRNA from miR-430. In this scenario (e.g. storage in a specific subcellular compartment), nanos1 reporter mRNA would be inaccessible to miRNAs in germ cells. To test this model, we introduced three copies of the miR-430 target site in the 3′UTR. Strikingly, this derivative of the nanos1 3′UTR was repressed by miR-430 in both PGCs and somatic cells (Fig.4H). Similarly, injection of high amounts of miR-204 inhibited the GFP expression of nanos1miR-204 mRNA in PGCs and somatic cells (data not shown). Taken together, these experiments indicate that the nanos1 3′UTR is accessible to miRNAs in PGCs and that the nanos1 3′UTR contains elements that allow the post-transcriptional activation in PGCs even in the presence of miR-430.
Figure 4. The nanos1 3′UTR allows protein expression in PGCs in the presence of miRNAs.

(A) Schematic of GFP-nanos1 3′UTR reporter constructs. Black bar indicates nanos1 3′UTR. Open box indicates miR-430 target site and gray box indicates miR-204 site. GFP reporter mRNA was injected at the one-cell stage, and GFP expression in PGCs was assayed at 26 hpf. To visualize PGCs, DsRed mRNA containing the nanos1 3′UTR was co-injected. GFP expression levels in PGCs and somatic cells are indicated on the right. N.D : not determined.(B–H) Expression of GFP reporter in gonad region (green) at 26 hpf. (B’–H’) Control DsRed expression in PGCs (red). Scale bar in H’shows 100μm.
The TDRD7 3′UTR is targeted by miR-430 and directs protein expression in PGCs
To determine if the regulation of germline genes by miR-430 is a general phenomenon, we asked whether miR-430 regulates the expression of other PGC-specific mRNAs. Using the ZFIN gene expression database (http://zfin.org), we found that zgc:56669, which encodes Tudor domain containing protein 7 (TDRD7), includes two GCACUU sequences in its 3′UTR. In situ hybridization showed that zgc:56669/TDRD7 is expressed maternally and restricted to PGCs, similar to nanos1 (Fig.5A-E and data not shown). Reporter assays revealed that the zgc:56669/TDRD7 3′UTR repressed protein synthesis in somatic cells, similar to the nanos1 3′UTR (Fig.5G). Two lines of evidence suggest that repression of GFP-zgc:56669/TDRD7 in somatic cells depends on miR-430. First, GFP expression of the TDRD7 reporter was higher in somatic cells that lack miR-430 (MZdicer mutants) than in wild-type embryos (Fig.5G and H). Second, mutations in the predicted target sites (Fig. 5F) abolished repression of the GFP reporter in somatic cells (Fig.5I and J). These results indicate that both the nanos1 and zgc:56669/TDRD7 3′UTRs mediate miR-430-induced repression in somatic cells.
Figure 5. The TDRD7 3′UTR is targeted by miR-430 in somatic cells and allows protein expression in PGCs.
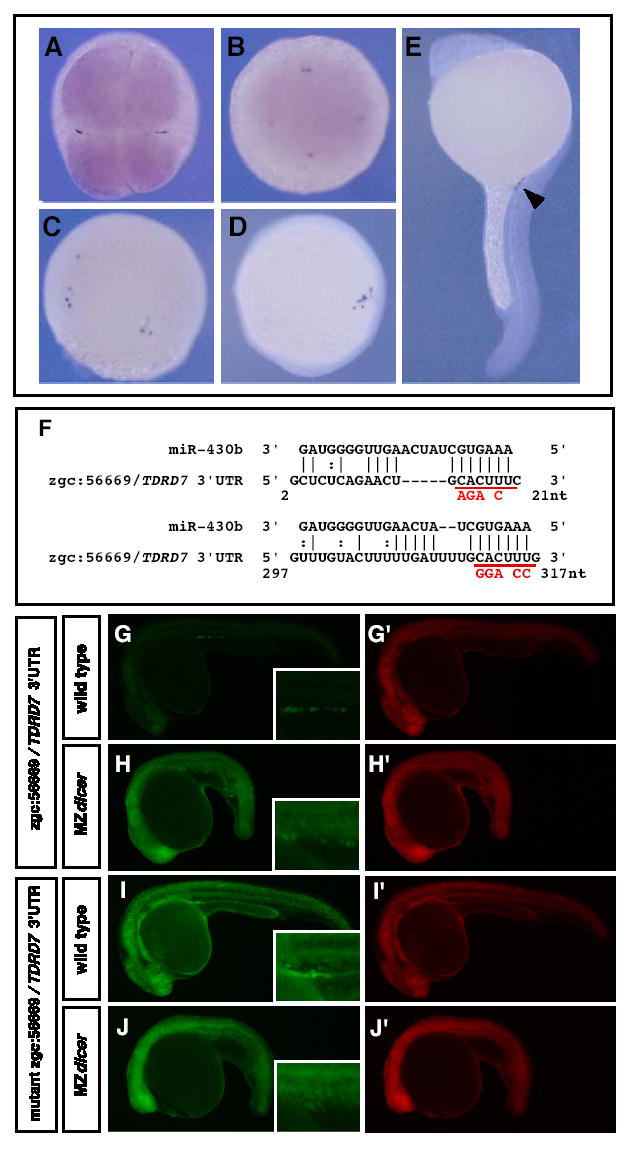
(A–D) In situ hybridization of zgc:56669/TDRD7 at (A) 4-cell, (B) sphere, (C) 80% epiboly, (D) 3-somite (E) 24 hpf (dark purple). Arrowhead indicates PGCs. A and B are animal pole views, whereas C, D, E are lateral views. (F) Putative base pairing of miR-430b with zgc:56669/TDRD7 3′UTR. Base-substitutions to disrupt the putative miR-430b binding are shown in red. The positions relative to the stop codon are shown. (G–I) GFP mRNA with wild-type or mutant zgc:56669/TDRD7 3′UTR were injected into wild-type (G, I) or MZdicer embryos (H, J) at the one-cell stage. GFP expression patterns at 26 hpf are shown. Inset shows enlarged view of the gonad region. (G’–J’) Control DsRed expression at 26 hpf.
Discussion
Our study indicates that miR-430 targets the 3′UTR of zebrafish nanos1 to induce mRNA deadenylation, mRNA degradation and translational repression in somatic cells (Fig.3). Conversely, the nanos1 3′UTR also includes cis-acting elements that allow activity in PGCs even in the presence of miRNA-mediated repression (Fig.4). Since another germline gene, TDRD7, is also targeted by miR-430, our results suggest that the differential effects of miR-430 in somatic cells and PGCs contribute to germline-specific gene expression.
While miR-430 is an important regulator of the nanos1 3′UTR, two observations suggest that there are additional mechanisms that regulate nanos1 expression. First, miR-430 is expressed at the onset of zygotic transcription (MBT) [9], whereas maternally provided GFP-nanos1 mRNA is posttranscriptionally repressed already prior to MBT [12]. Second, the degradation of endogenous nanos1 mRNA still occurs in MZdicer mutants (data not shown). Therefore we propose that miR-430 contributes to, but is not the sole determinant of the soma-specific decay and translational repression of nanos1 mRNA. Indeed, studies in Drosophila have shown that nanos mRNA is regulated by multiple post-transcriptional mechanisms including deadenylation, mRNA decay and translational repression [6, 31–35].
Previous reporter studies have shown that miR-430 target mRNAs are equally susceptible to repression in somatic cells and PGCs [9, 19]. The results presented here identify a novel class of miR-430 targets that is differentially regulated between soma and germline. In the case of nanos1, differential repression is due to cis-acting elements in the 3′UTR. The exact role of these sequences is not known, but our study excludes several simple models. First, local masking of the miR-430 binding site is unlikely to be responsible for PGC-specific activation, because the nanos1 3′UTR can promote protein synthesis in PGCs independent of the sequence and location of the miRNA target site (Fig.4E, F and G). Second, it is unlikely that the nanos1 mRNA is sequestered from miR-430, because extra copies of the miR-430 target site make the nanos1 reporter susceptible to repression in PGCs (Fig.4H). Excluding these models, we speculate that PGCs promote the expression of nanos1 and other germline-specific mRNAs not by inactivation of the miRNA or its associated machinery but by recruiting other factors to the 3′UTR that increase mRNA stability or translation.
In addition to revealing a role for microRNAs in soma versus germline gene expression, our results have wider implications for the regulation of mRNAs by miRNAs. We found that miRNAs can be effective regulators of a target mRNA in one tissue but ineffective in another tissue. Hence, the presence of a miRNA target site and repression in one tissue (e.g. somatic cells) does not necessarily result in repression in another tissue (e.g. PGCs). Analogously, it has been shown that some miRNA targets have differential susceptibilities under stress and normal conditions [36]. Hence, miRNA-mediated regulation is conditional on tissue-specific or cell-state specific factors. Recent studies have also shown that some predicted miRNA targets are expressed at high levels in cells that express the cognate miRNA [19, 29, 37, 38]. These observations and our results suggest that there are not only “anti-targets”, which have evolved 3′UTRs that lack miRNA target sites [37, 38], but also “resistant targets”, which have evolved mechanisms to counteract the effects of miRNAs.
Supplementary Material
Acknowledgments
We thank members of our labs, especially T. Takasaki and K. Fukumura, for valuable discussions, the Yasuda lab members for help during the initial phase of the project, and H. Knaut for discussions and critical reading of the manuscript. This work was supported by Grants-in-Aid from MEXT and JSPS, and also supported in part by The Asahi Glass Foundation to K.I. Y.M was supported by JSPS Research Fellowships. A.J.G. was supported by EMBO and is currently supported by a Human Frontier Science Program fellowship. A.F.S. was an Irma T. Hirschl Trust Career Scientist and an established Investigator of the American Heart Association. This work was also supported by grants from the NIH (A.F.S).
References
- 1.Wylie C. Germ cells. Curr Opin Genet Dev. 2000;10:410–413. doi: 10.1016/s0959-437x(00)00105-2. [DOI] [PubMed] [Google Scholar]
- 2.Kobayashi S, Yamada M, Asaoka M, Kitamura T. Essential role of the posterior morphogen nanos for germline development in Drosophila. Nature. 1996;380:708–711. doi: 10.1038/380708a0. [DOI] [PubMed] [Google Scholar]
- 3.Subramaniam K, Seydoux G. nos-1 and nos-2, two genes related to Drosophila nanos, regulate primordial germ cell development and survival in Caenorhabditis elegans. Development. 1999;126:4861–4871. doi: 10.1242/dev.126.21.4861. [DOI] [PubMed] [Google Scholar]
- 4.Koprunner M, Thisse C, Thisse B, Raz E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes Dev. 2001;15:2877–2885. doi: 10.1101/gad.212401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Tsuda M, Sasaoka Y, Kiso M, Abe K, Haraguchi S, Kobayashi S, Saga Y. Conserved role of nanos proteins in germ cell development. Science. 2003;301:1239–1241. doi: 10.1126/science.1085222. [DOI] [PubMed] [Google Scholar]
- 6.Gavis ER, Lehmann R. Translational regulation of nanos by RNA localization. Nature. 1994;369:315–318. doi: 10.1038/369315a0. [DOI] [PubMed] [Google Scholar]
- 7.Bashirullah A, Halsell SR, Cooperstock RL, Kloc M, Karaiskakis A, Fisher WW, Fu W, Hamilton JK, Etkin LD, Lipshitz HD. Joint action of two RNA degradation pathways controls the timing of maternal transcript elimination at the midblastula transition in Drosophila melanogaster. EMBO J. 1999;18:2610–2620. doi: 10.1093/emboj/18.9.2610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.D’Agostino I, Merritt C, Chen PL, Seydoux G, Subramaniam K. Translational repression restricts expression of the C. elegans Nanos homolog NOS-2 to the embryonic germline. Dev Biol. 2006;292:244–252. doi: 10.1016/j.ydbio.2005.11.046. [DOI] [PubMed] [Google Scholar]
- 9.Giraldez AJ, Cinalli RM, Glasner ME, Enright AJ, Thomson JM, Baskerville S, Hammond SM, Bartel DP, Schier AF. MicroRNAs regulate brain morphogenesis in zebrafish. Science. 2005;308:833–838. doi: 10.1126/science.1109020. [DOI] [PubMed] [Google Scholar]
- 10.Hashimoto Y, Maegawa S, Nagai T, Yamaha E, Suzuki H, Yasuda K, Inoue K. Localized maternal factors are required for zebrafish germ cell formation. Dev Biol. 2004;268:152–161. doi: 10.1016/j.ydbio.2003.12.013. [DOI] [PubMed] [Google Scholar]
- 11.Theusch EV, Brown KJ, Pelegri F. Separate pathways of RNA recruitment lead to the compartmentalization of the zebrafish germ plasm. Dev Biol. 2006;292:129–141. doi: 10.1016/j.ydbio.2005.12.045. [DOI] [PubMed] [Google Scholar]
- 12.Blaser H, Eisenbeiss S, Neumann M, Reichman-Fried M, Thisse B, Thisse C, Raz E. Transition from non-motile behaviour to directed migration during early PGC development in zebrafish. J Cell Sci. 2005;118:4027–4038. doi: 10.1242/jcs.02522. [DOI] [PubMed] [Google Scholar]
- 13.Richter JD. Cytoplasmic polyadenylation in development and beyond. Microbiol Mol Biol Rev. 1999;63:446–456. doi: 10.1128/mmbr.63.2.446-456.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wilkie GS, Dickson KS, Gray NK. Regulation of mRNA translation by 5′- and 3′-UTR-binding factors. Trends Biochem Sci. 2003;28:182–188. doi: 10.1016/S0968-0004(03)00051-3. [DOI] [PubMed] [Google Scholar]
- 15.de Moor CH, Meijer H, Lissenden S. Mechanisms of translational control by the 3′ UTR in development and differentiation. Semin Cell Dev Biol. 2005;16:49–58. doi: 10.1016/j.semcdb.2004.11.007. [DOI] [PubMed] [Google Scholar]
- 16.Wolke U, Weidinger G, Koprunner M, Raz E. Multiple levels of posttranscriptional control lead to germ line- specific gene expression in the zebrafish. Curr Biol. 2002;12:289–294. doi: 10.1016/s0960-9822(02)00679-6. [DOI] [PubMed] [Google Scholar]
- 17.Schwartz DC, Parker R. Mutations in translation initiation factors lead to increased rates of deadenylation and decapping of mRNAs in Saccharomyces cerevisiae. Mol Cell Biol. 1999;19:5247–5256. doi: 10.1128/mcb.19.8.5247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bergamini G, Preiss T, Hentze MW. Picornavirus IRESes and the poly(A) tail jointly promote cap-independent translation in a mammalian cell-free system. RNA. 2000;6:1781–1790. doi: 10.1017/s1355838200001679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Giraldez AJ, Mishima Y, Rihel J, Grocock RJ, Van Dongen S, Inoue K, Enright AJ, Schier AF. Zebrafish MiR-430 promotes deadenylation and clearance of maternal mRNAs. Science. 2006;312:75–79. doi: 10.1126/science.1122689. [DOI] [PubMed] [Google Scholar]
- 20.Wu L, Fan J, Belasco JG. MicroRNAs direct rapid deadenylation of mRNA. Proc Natl Acad Sci U S A. 2006;103:4034–4039. doi: 10.1073/pnas.0510928103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Behm-Ansmant I, Rehwinkel J, Doerks T, Stark A, Bork P, Izaurralde E. mRNA degradation by miRNAs and GW182 requires both CCR4:NOT deadenylase and DCP1:DCP2 decapping complexes. Genes Dev. 2006;20:1885–1898. doi: 10.1101/gad.1424106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- 23.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 24.Du T, Zamore PD. microPrimer: the biogenesis and function of microRNA. Development. 2005;132:4645–4652. doi: 10.1242/dev.02070. [DOI] [PubMed] [Google Scholar]
- 25.Brennecke J, Stark A, Russell RB, Cohen SM. Principles of microRNA-target recognition. PLoS Biol. 2005;3:e85. doi: 10.1371/journal.pbio.0030085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Doench JG, Sharp PA. Specificity of microRNA target selection in translational repression. Genes Dev. 2004;18:504–511. doi: 10.1101/gad.1184404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kloosterman WP, Wienholds E, Ketting RF, Plasterk RH. Substrate requirements for let-7 function in the developing zebrafish embryo. Nucleic Acids Res. 2004;32:6284–6291. doi: 10.1093/nar/gkh968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lai EC. Micro RNAs are complementary to 3′ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat Genet. 2002;30:363–364. doi: 10.1038/ng865. [DOI] [PubMed] [Google Scholar]
- 29.Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, Bartel DP, Linsley PS, Johnson JM. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature. 2005;433:769–773. doi: 10.1038/nature03315. [DOI] [PubMed] [Google Scholar]
- 30.Bagga S, Bracht J, Hunter S, Massirer K, Holtz J, Eachus R, Pasquinelli AE. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell. 2005;122:553–563. doi: 10.1016/j.cell.2005.07.031. [DOI] [PubMed] [Google Scholar]
- 31.Dahanukar A, Walker JA, Wharton RP. Smaug, a novel RNA-binding protein that operates a translational switch in Drosophila. Mol Cell. 1999;4:209–218. doi: 10.1016/s1097-2765(00)80368-8. [DOI] [PubMed] [Google Scholar]
- 32.Smibert CA, Lie YS, Shillinglaw W, Henzel WJ, Macdonald PM. Smaug, a novel and conserved protein, contributes to repression of nanos mRNA translation in vitro. RNA. 1999;5:1535–1547. doi: 10.1017/s1355838299991392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Forrest KM, Clark IE, Jain RA, Gavis ER. Temporal complexity within a translational control element in the nanos mRNA. Development. 2004;131:5849–5857. doi: 10.1242/dev.01460. [DOI] [PubMed] [Google Scholar]
- 34.Kalifa Y, Huang T, Rosen LN, Chatterjee S, Gavis ER. Glorund, a Drosophila hnRNP F/H Homolog, Is an Ovarian Repressor of nanos Translation. Dev Cell. 2006;10:291–301. doi: 10.1016/j.devcel.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 35.Jeske M, Meyer S, Temme C, Freudenreich D, Wahle E. Rapid ATP-dependent deadenylation of nanos mRNA in a cell-free system from Drosophila embryos. J Biol Chem. 2006;281:25124–25133. doi: 10.1074/jbc.M604802200. [DOI] [PubMed] [Google Scholar]
- 36.Bhattacharyya SN, Habermacher R, Martine U, Closs EI, Filipowicz W. Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell. 2006;125:1111–1124. doi: 10.1016/j.cell.2006.04.031. [DOI] [PubMed] [Google Scholar]
- 37.Stark A, Brennecke J, Bushati N, Russell RB, Cohen SM. Animal MicroRNAs confer robustness to gene expression and have a significant impact on 3′UTR evolution. Cell. 2005;123:1133–1146. doi: 10.1016/j.cell.2005.11.023. [DOI] [PubMed] [Google Scholar]
- 38.Farh KK, Grimson A, Jan C, Lewis BP, Johnston WK, Lim LP, Burge CB, Bartel DP. The widespread impact of mammalian MicroRNAs on mRNA repression and evolution. Science. 2005;310:1817–1821. doi: 10.1126/science.1121158. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.