

Abstract
Background
Cell culture technology has spread prolifically within a century, a variety of culture media has been designed. This review goes through the history, characteristics and current issues of animal‐cell culture media.
Methods
A literature search was performed on PubMed and Google Scholar between 1880 and May 2016 using appropriate keywords.
Results
At the dawn of cell culture technology, the major components of media were naturally derived products such as serum. The field then gradually shifted to the use of chemical‐based synthetic media because naturally derived ingredients have their disadvantages such as large batch‐to‐batch variation. Today, industrially important cells can be cultured in synthetic media. Nevertheless, the combinations and concentrations of the components in these media remain to be optimized. In addition, serum‐containing media are still in general use in the field of basic research. In the fields of assisted reproductive technologies and regenerative medicine, some of the medium components are naturally derived in nearly all instances.
Conclusions
Further improvements of culture media are desirable, which will certainly contribute to a reduction in the experimental variation, enhance productivity among biopharmaceuticals, improve treatment outcomes of assisted reproductive technologies, and facilitate implementation and popularization of regenerative medicine.
Keywords: cell culture technique, cell proliferation, culture media, cultured cells, serum
1. Introduction
The influence of cell culture technology on human society has been immeasurable. Progress in biology in recent years, for example, has depended heavily on cell culture technology.1 In addition, cell culture‐based practical technologies have been developed in various areas, including the assessment of the efficacy and toxicity of new drugs, manufacture of vaccines and biopharmaceuticals, and assisted reproductive technology. As the reprogramming of somatic cells became technically feasible recently, researchers around the world are fiercely competing for leadership in the advances of regenerative medicine. In this area likewise, cell culture technology is regarded as a foundation for further development and popularization.
No one probably would argue against the claim that a culture medium is the most important factor in cell culture technology. A medium supports cell survival and proliferation, as well as cellular functions, meaning that the quality of the medium directly affects the research results, the biopharmaceutical production rate, and treatment outcomes of assisted reproductive technology. It is essential, therefore, for investigators who are working with cell cultures to select an appropriate medium that is suitable for their aims. In some cases, researchers should modify a medium themselves. In addition, when facing problems, researchers have to know the properties of the medium in order to identify the cause of any problem with their experiments.
At present, synthetic media can be classified into several groups, based on the type of supplements added; for example, serum‐containing media, serum‐free media, protein‐free media, and chemically defined media (Tables 1 and 2). Serum‐containing media naturally contain various serum‐derived substances, which make the medium composition unclear and whose concentrations can fluctuate from batch to batch. This situation makes the culture results less reproducible and poses a risk of microbial contamination. Serum‐containing media, however, can be designed easily and be used effectively for a variety of cell types because serum includes a lot of active substances that are necessary for the survival and growth of animal cells.2 Serum‐free media, in contrast, have a defined composition, resulting in a high reproducibility of results, and the cultivation process can be validated. In addition, target cells can be grown selectively in an intermingled cell population if the culture conditions are configured to benefit them. Among the serum‐free media, subgroups of protein‐free media (which do not contain any protein at all) and chemically defined media (which do not contain any undefined ingredient) provide additional stability and reproducibility for culture systems, facilitating the identification of the cellular secretions and reducing the risk of microbial contamination. However, the serum‐free media are difficult to design: only specific cell types have been cultivated this way to date.3
Table 1.
Categories of animal‐cell culture media
| Category | Definition | Type | Example |
|---|---|---|---|
| Natural media | Consisting of natural biological substances, such as plasma, serum, and embryo extract | Coagulant or clots | Plasma separated from heparinized blood, serum, and fibrinogen |
| Tissue extracts | Extracts of chicken embryos, liver, and spleen and bone marrow extract | ||
| Biological fluids | Plasma, serum, lymph, amniotic fluid, and pleural fluid | ||
| Synthetic media | Composed of a basal medium and supplements, such as serum, growth factors, and hormones | Serum‐containing media | Human, bovine, equine, or other serum is used as a supplement |
| Serum‐free media | Crude protein fractions, such as bovine serum albumin or α‐ or β‐globulin, are used as supplements | ||
| Xeno‐free media | Human‐source components, such as human serum albumin, are used as supplements but animal components are not allowed as supplements | ||
| Protein‐free media | Undefined components, such as peptide fractions (protein hydrolysates) are used as supplements | ||
| Chemically defined media | Undefined components, such as crude protein fractions, hydrolysates, and tissue extracts, are not appropriate as supplements, but highly purified components, such as recombinant proteins are appropriate supplements |
Table 2.
Types and characteristics of basal media
| Category | Name (author, year) | Features |
|---|---|---|
| Connaught Medical Research Laboratories (CMRL) media | Medium 199 (Morgan et al. 1950) | Developed in order to cultivate chicken embryonic cells under protein‐free conditions, it is prepared by the sequential addition of amino acids, vitamins (including fat‐soluble vitamins), and nucleic‐acid precursors. Its composition is extremely complex because the components that are thought to be necessary on theoretical grounds, including inactive components, are added to the medium. Often used for organ culture |
| CMRL1066 (Parker et al. 1957) | Created by amending Medium 199 in order to culture mouse L cells under protein‐free conditions, the modifications included increased levels of reducing substances (cysteine, glutathione, and ascorbic acid), the elimination of fat‐soluble vitamins, changes in the types of nucleic‐acid precursors, and the addition of coenzymes | |
| Eagle media | Basal medium Eagle (BME) (Eagle 1955) | Supplemented with the minimal components that are necessary for mouse L cells and human HeLa cells to reach the index of proliferative capacity and including 13 amino acids and eight vitamins, it is unsuitable for cells whose cultures require many components because of its simple composition |
| Minimum essential medium (MEM) (Eagle 1959) | BME modified according to the cellular need for amino acids, so the concentrations of the majority of amino acids are twofold, as compared to BME. Non‐essential amino acids, which cells can biosynthesize, were not included in the original MEM formulation; however, researchers can add non‐essential amino acids to reduce the biosynthetic load | |
| Dulbecco's modified MEM (DMEM) (Dulbecco and Freeman 1959) | Modified to have fourfold the concentrations of amino acids and vitamins that are present in BME and developed to study the plaque‐forming ability of the polyoma virus in mouse embryonic cells. Various modifications have been made since, with supplementation, for example, of the non‐essential amino acids, glycine and serine, iron, and pyruvate. The glucose concentration also can be increased to 25 mmol L−1 in order to accommodate cells with high nutritional requirements. In the event that pH changes are suspected due to metabolites, sodium bicarbonate is doubled in concentration, equilibrated, and then used at 10% CO2 | |
| α‐MEM (Stanners et al. 1971) | MEM modified for hybrid‐cell‐line research on mice and hamsters, its composition is MEM supplemented with non‐essential amino acids and vitamins (ascorbic acid, biotin, and cyanocobalamin), pyruvate, lipoic acid, and nucleosides | |
| Iscove's modified DMEM (IMDM) (Iscove and Melchers 1978) | Supplemented with several non‐essential amino acids and vitamins that are not present in DMEM (cyanocobalamin and biotin) and additional selenite, pyruvate, and HEPES. Transferrin, bovine serum albumin, and soybean lipids are added as serum substitutes. With its high concentrations of amino acids and vitamins, IMDM is suitable for high‐density cultures and cultures of rapidly proliferating cells | |
| Tissue Culture Section of the National Cancer Institute (NCTC) media | NCTC109 (McQuilkin et al. 1957) | Developed to culture L cells under protein‐free conditions, its amino acid composition is based on the results of a componential analysis of the compounds that have been ultrafiltered from horse serum and chicken embryonic‐tissue extract. Its composition is quite complex, with not only coenzymes, nucleobases, and reducing agents, but a wealth of vitamins added, as well (ie, A, C, D, E, and K, in addition to the B‐group vitamins). Cysteine was included in the original composition, but after it was found to negatively affect cells, a version was developed from which cysteine was removed (NCTC135) |
| Ham media | Ham's F‐10 (Ham 1963) | Enables colony formation by a single Chinese hamster ovary (CHO) cell under serum‐free conditions, developed by adding two kinds of purified serum proteins (serum albumin and fetuin), instead of serum, and by examining in detail the types and concentrations of amino acids and trace elements This medium was the first to contain the trace elements, copper and zinc (iron having been included in other media already). The CHO cells have an inferior proliferative capacity in this medium alone, compared to the one with serum added. Furthermore, culture of the cell lines other than CHO necessitates the addition of serum |
| Ham's F‐12 (Ham 1965) | The serum albumin and fetuin (used in Ham's F‐10) are replaced by two compounds with definite chemical composition: linoleic acid and putrescine, which enables colony formation by a single CHO cell under protein‐free conditions. Often cited as the world's first chemically defined medium, the levels of several amino acids are higher than in Ham's F‐10, while those of the vitamins (except choline and inositol) and potassium phosphate are reduced. Its composition must be modified (eg, by reducing the zinc concentration) for a protein‐free culture of cells other than CHO cells. MCDB301, in which 20 trace elements were added, was later developed after it became obvious that trace elements contaminating water or raw materials are necessary for a protein‐free culture of CHO cells in Ham's F‐12. | |
| Kaighn's modified Ham's F‐12 (Ham's F‐12K) (Kaighn 1974) | The concentrations of the amino acids, pyruvate, biotin, calcium, magnesium, putrescine, and phenol red are increased with respect to those in Ham's F‐12, among other compositional modifications, in order to support the proliferation and differentiation of primary cultured cells | |
| Roswell Park Memorial Institute (RPMI) media | RPMI 1640 (Moore et al. 1966) | Based on the 5A Medium (developed by McCoy et al. 1959) and modified for the long‐term culture of peripheral blood lymphocytes, it is characterized by low levels of calcium and magnesium and high levels of phosphate. Multiple media were developed on the path to this medium (eg, RPMI 1629, 1630, and 1634). It is widely used as a medium for suspension cultures; for example, of white blood cells, lymphocytes, and hybridomas |
| Molecular, Cellular, and Development Biology (MCDB) media |
For example, MCDB202 (McKeehan et al. 1976) MCDB301 (Hamilton et al. 1977) MCDB153 (Peehl et al. 1980) MCDB110 (Bettger et al. 1981) MCDB402 (Shipley et al. 1983) MCDB170 (Hammond et al. 1984) MCDB131 (Knedler et al. 1987) |
A series of Ham's F‐12‐based media that was developed in order to grow specific cell types in serum‐free culture. The composition of each MCDB medium is optimized to promote the growth of a specific cell type. In particular, MCDB202 is a medium for the serum‐free culture of chicken embryo fibroblasts; MCDB301 is for the serum‐free culture of CHO cells; MCDB153 is for human keratinocytes; MCDB110 is for human fibroblasts; MCDB402 is for mouse fibroblasts; MCDB170 is for the mammary epithelium; and MCDB131 is a medium for the serum‐free culture of human vascular endothelial cells |
| Mixed media | DMEM/F‐12 (Barnes and Sato 1979) | Constitutes a 50:50 mixture of component‐rich Ham's F12 medium and the nutrient‐rich DMEM medium and compatible with the requirements of a variety of cell types, it is most often used as a basal medium for serum‐free culture |
| RPMI 1640/DMEM/F‐12 (RDF) (Murakami 1984) | Developed for the serum‐free culture of hybridomas, it is a mixture of RPMI 1640, DMEM, and Ham's F‐12 at 2:1:1 and is typically used as a serum‐free medium with added insulin, transferrin, ethanolamine, and selenite | |
| Other media | Waymouth's MB752/1 (Waymouth 1959) | Developed with as simple a composition as feasible, so that mouse L929 cells could be cultured without the addition of serum and other proteins, it is composed of a total of 40 components, including glucose, inorganic salts, amino acids, vitamins, purine bases, and hypoxanthine. It is characterized by high concentrations of glucose, histidine, lysine, glutamine, choline, and thiamine |
| Trowell's T‐8 (Trowell 1959) | Designed for the long‐term culture of adult rat liver epithelial cells. With its comparatively simple composition, it contains no non‐essential amino acid and hardly any vitamins, but is characterized by high glucose and insulin concentrations. It is used for short‐term organ culture | |
| Leibovitz's L‐15 (Leibovitz 1963) | The buffering capacity is mediated by phosphates and free basic amino acids instead of sodium bicarbonate, so that a culture's pH is maintained in ambient air without the use of a CO2 incubator. Instead of glucose, pyruvate (and galactose) is added at a high concentration in order to control pH drops due to the lactic acid that is produced during glucose metabolism and to promote the release of CO2 from the respiratory chain. Another major characteristic is that each of its constituent amino acids is added at approximately its respective maximal concentration. Once used for cell and tissue transport and primary cultures, its popularity has declined as researchers began to use HEPES as a buffering agent and as they realized, moreover, that a certain amount of sodium bicarbonate is necessary for optimal cell proliferation | |
| Fischer's Medium (Fischer and Sartorelli 1964) | Contains a high concentration of folate because this medium was developed by using folate‐dependent L5178Y lymphoma cells |
HEPES, 4‐(2‐hydroxyethyl)‐1‐piperazineethanesulfonic acid.
At present, formulations of popular media for cell culture are available commercially. Thus, there are investigators who are using culture media without understanding their details and background, particularly regarding the rationale for their development, the exact ingredients, as well as the cell types that these media are suitable for. This review article briefly describes the history of the development of animal‐cell culture media, with comments on the types of media in general use today regarding their characteristics, roles of the medium components, and pitfalls or problems with their use.
2. History of Cell Culture Media
2.1. Dawn of cultivation experiments (1882–1907)
In 1882, Sydney Ringer developed Ringer's solution, a balanced salt solution of a composition that is close to that of bodily fluids, and successfully kept frog hearts beating after dissection and removal from the body.4, 5 This is said to be the first instance of in vitro cultivation of animal tissue. Balanced salt solutions were developed one after another in the wake of Ringer's report, including Locke's solution,6 Tyrode's solution,7 the Krebs–Ringer bicarbonate solution,8 Gey's solution,9 Earle's solution,10 and Hanks' solution.11 The composition of these balanced salt solutions is simple and includes only inorganic salts, sometimes with glucose added as a nutrient. Nonetheless, their pH, osmotic pressure, and inorganic salt concentrations were calibrated to physiological conditions and these solutions can be used successfully to keep tissues and cells outside the body alive for short periods, generally up to a few days.
After the success of Ringer's solution, researchers began to pay attention to cells in culture devices and tried to maintain the cells. Nonetheless, the cells usually did not survive and rarely showed mitotic figures.12, 13, 14 In 1907, however, Ross G. Harrison successfully monitored an apparent outgrowth of nerve fibers of a frog for several weeks in lymph fluid that had been freshly drawn from the lymph sacs of an adult frog.15 This experiment is considered to be the beginning of animal cell cultivation.
2.2. Use of natural media (1907‐)
Alexis Carrel was a French surgeon and biologist who received the Nobel Prize in Physiology and Medicine in 1912 for his research on the vascular suture and transplantation of blood vessels and organs. He contributed greatly to tissue culture technology by devising a prototype of the cell culture flask that is used widely today and by establishing the aseptic manipulation technique.16
The first success of animal cell culture by Harrison inspired Carrel to send Montrose T. Burrows to work under Harrison's supervision in 1909. There, Burrows found that lymph is unsuitable for the cultivation of cells from warm‐blooded animals and used plasma instead. Thereafter, blood plasma had become a major culture medium for a variety of animal cells. He successfully cultivated chicken embryonic cells by using chicken blood plasma, which is readily available,17 and later successfully cultivated mammalian cells as well.18 In 1912, Carrel demonstrated that the long‐term cultivation of the cells that have been obtained from the connective tissues of chick fetuses is possible (for several months) with a periodic exchange of the medium.19 In 1913, he discovered that adding embryonic extract to blood plasma can dramatically increase cellular proliferation and extend the culture period of fibroblasts from the chick embryo heart.20, 21 Meanwhile, because the composition of the lymph, plasma, and embryonic extract was unknown, it became a new scientific inquiry regarding which of their components affected the survival and growth of animal tissues and cells. This situation led to a period when researchers attempted to identify the growth‐promoting substances within these ingredients of natural origin and to replace them with ingredients of definite composition.
Carrel is widely believed to be the first person in the world to successfully culture mammalian somatic cells, but the Biographical Memoirs of the National Academy of Sciences contends that the cultivation of guinea pig bone marrow specimens in 1908 by Margaret Reed, the future Mrs. Warren H. Lewis, precedes his success.22
2.3. Endeavors toward synthetic media (1911‐)
Margaret R. Lewis and Warren H. Lewis (1911) demonstrated that the Locke–Lewis solution—which is modified Locke's solution that additionally contains amino acids, bouillon, and glucose (or maltose)—is more effective for chick embryo cell cultivation than simple balanced salt solutions.23, 24 They reported that glucose is especially important: if the concentration of glucose is not sufficient in the medium, the chick embryo cells show vacuolar degeneration and die within a few days.25 In contrast, in their search for the active ingredients in embryonic extract, it was ascertained that the active substance is in the protein fraction26 and that the partially hydrolyzed proteins effectively promote the cell growth of chick embryo fibroblasts.27 Those researchers also confirmed the growth‐promoting activity of amino acids28 and glutathione for chick embryo fibroblasts.29 They hypothesized that glutathione is required for the control of the redox environment during cell cultivation. Carrel's medium was supplemented with several natural products, such as casein digests (thymus‐derived) nucleic acids, liver ash, and hemoglobin. Thus, their research did not lead to the molecular identification of any substance that is essential for cultured cells. Joilannes P. M. Vogelaar and Eleanor Erlichman, in contrast, confirmed the effectiveness of two hormones (insulin and thyroxine) by successfully cultivating human fibroblasts for >3 months in a medium that consisted of Ringer's solution with these hormones, along with glucose, cysteine, hemin, peptone, and blood plasma.30 Researchers continued the efforts to specify the composition of media thereafter, including the development of Baker's medium, which contains vitamin A, ascorbic acid, vitamin B1, vitamin B2, glutathione, and blood plasma.31 Nevertheless, this approach did not lead to culture media that did not include natural products, such as blood serum or plasma.
2.4. Birth of established cell lines (1940‐)
It is rare for healthy somatic cells that are derived from animals to acquire unlimited proliferative capacity during cultivation. They typically stop growing after a certain number of divisions (ie, the Hayflick limit).32 As a result, researchers would conduct studies by using cells that had been sampled from animals in each experiment. In 1940, Wilton R. Earle et al. used carcinogens to successfully create immortal mouse fibroblasts (L cells),10 revealing that proliferation from a single cell is possible.33 In 1951, George O. Gey and his coworkers created an infinitely proliferating human cell line from a tissue of a patient with uterine cervical cancer (HeLa cells).34 Due to the emergence of these established cell lines, the sampling of cells from the tissues of animals in each experiment became unnecessary, enabling researchers worldwide to perform assays by using the same homogenous population of cells. This state of affairs made it easier to examine and to precisely quantify the subtle differences in the effects of culture media on cells. Thus, the development of culture media advanced rapidly as a result.
2.5. Establishment of basal media and research into protein‐free media (1946‐)
Baker's medium and the other media that had been developed up to this point contained naturally derived components of unknown composition, including plasma, serum, bouillon, peptone, and tissue extracts. In order to find the crucial components in those natural materials and to develop defined media that are comparably efficient in the cultivation of cells, relative to the media containing natural ingredients, two main strategies were undertaken. The first strategy was to use dialyzed serum for the support of cells at minimum levels and to add defined components to maximize the proliferation of cells. The second strategy was not dependent on serum, or even proteins at all, and involved the formulation of media exclusively from definitive components.
Fischer was a pioneer of the first strategy. He dialyzed blood plasma to remove the low‐molecular‐weight fraction. Culture media that were supplemented with dialyzed blood plasma could sustain cells only for a short period, indicating that the low‐molecular‐weight fraction was essential for the survival of cells. Then, he discovered that the amino acids are the key substance in the low‐molecular‐weight fraction.35, 36 This creation of dialyzed media, along with the established cell lines, made it possible to determine, in a systemic way, whether the cells require low‐molecular‐weight substances under culture conditions. For example, in 1955, Harry Eagle studied the minimum necessary amounts of low‐molecular‐weight components that are required by mouse L cells and HeLa cells by using a balanced salt solution with the addition of dialyzed serum, on the basis of Fischer's method.37, 38 He found that 13 amino acids and eight vitamins are necessary.39 Going on to study the amino acid requirements of a variety of cells, Eagle then developed the minimum essential medium (MEM), which is composed of the minimal essential components that he identified (glucose, six inorganic salts, 13 amino acids, eight water‐soluble vitamins, and dialyzed serum).40 Renato Dulbecco and Marguerite Vogt,41 Clifford P. Stanners et al.,42 and Norman N. Iscove and Fritz Melchers43 made improvements to MEM, according to the desired cell type and culture purpose (Table 2). Later, using dialyzed serum that had been prepared in accordance with Fischer's method, Thomas A. McCoy et al. confirmed that carcinosarcoma cells require pyruvate.44 At this time, the Roswell Park Memorial Institute (RPMI) modified the 5A medium that had been developed by McCoy,45 in terms of its calcium and magnesium concentrations. The result then was reported as the RPMI 1640 medium for use in lymphocyte cultivation.46, 47
The other strategy—the total omission of serum and proteins—is thought to have begun in 1946, with full‐scale research by Philip R. White. He developed a chemically defined medium that was composed of glucose, inorganic salts, amino acids, iron, vitamins, and glutathione, with no protein at all. Using this medium, he successfully cultivated chick embryo‐derived fibroblasts and cardiac muscle cells for ~2 months.48 Other researchers were unable to repeat his findings and argued that White's Medium still needed 10%‐20% serum for the reported results.49 In 1950, Connaught Medical Research Laboratories (CMRL), led by Raymond C. Parker, developed Medium 199, which includes fat‐soluble vitamins, cholesterol, and nucleic acid precursors, in addition to the ingredients of White's Medium (Table 2). Instead of adding serum or proteins, Parker's team used the strategy of adding as many low‐molecular‐weight substances as possible, according to what was considered theoretically necessary for cell culture.50 Using this medium, they cultured chick embryo‐derived cells for 3‐4 weeks. Serum and embryonic extract were still necessary for the primary culture. Other researchers also reported the necessity of a small amount of serum in Medium 199 for cell proliferation.37 The team at CMRL thereafter developed a chemically defined medium, CMRL1066, using mouse L cells and their proliferation as an indicator of the effectiveness of the medium. This CMRL1066 medium, consisting of 58 components, was developed through several modifications, such as increasing the levels of reducing substances (cysteine, glutathione, and ascorbic acid), removing fat‐soluble vitamins, changing the nucleic acid precursors, and adding coenzymes (Table 2).51, 52 In contrast, during the same period, the National Cancer Institute's Tissue Culture Section (NCTC), led by Wilton R. Earle, developed a chemically defined medium that was composed of 68 ingredients, named NCTC109 (Table 2).53 These two media were difficult to prepare because of their complex composition. Thereafter, Charity Waymouth developed the MB 752/1 medium (Table 2), which is composed of 40 ingredients (glucose, inorganic salts, amino acids, vitamins, purine bases, hypoxanthine, and glutathione). This was the simplest‐possible chemically defined medium at the time.54 CMRL1066, NCTC109, and MB 752/1, however, all were designed for the best growth of mouse L cells. Therefore, they were not necessarily suitable for the serum‐free culture of other cell types. In addition, cell cloning was not possible in those protein‐free media. Waymouth hypothesized that the proteins that are produced by cells are necessary for proliferation under protein‐free conditions: the so‐called “auto/paracrine effect.” It was in this context that Richard G. Ham developed Ham's F‐10 medium in 1963 (Table 2). With two kinds of serum protein fractions (albumin and fetuin) instead of serum, he successfully made a single Chinese hamster ovary (CHO) cell form a colony under serum‐free conditions.53 The composition of this medium, however, was undefined because it contained serum protein fractions and the proliferative capacity it conferred fell short of that induced by serum‐containing media. There was also a challenge: serum supplementation remained necessary for the cultivation of cell types other than CHO cells. In order to solve these issues, Ham replaced the albumin and fetuin of biological origin with low‐molecular‐weight substances—linoleic acid and putrescine55, 56—and developed Ham's F‐12, a completely synthetic medium of definite composition. During this refinement of the medium, the concentration of each ingredient in Ham's F‐10 was reviewed and modified (Table 2).57 With the reduced amount of zinc, this medium supported colony formation by mouse L cells, but protein‐free cultivation of other cells remained difficult.58, 59 It became clear in later studies that trace elements contaminating the water or raw materials they used are necessary for the protein‐free cultivation of CHO cells in Ham's F‐12 medium, leading to the development of MCDB301, a medium that is supplemented with 20 trace elements (Table 2).60 Toshiko Takaoka and Hajim Katsuta successfully cultured various kinds of cells long‐term in a simple medium that did not contain proteins or lipids;61 however, only a few cell types can adapt to protein‐ and lipid‐free conditions and the fatty‐acid composition of the cells thus cultured differs greatly from the cells cultured in serum‐containing media.62
In the field of life sciences, the “reductive approach” and the “synthetic approach” are often used for elucidating a complicated biological process. The reductive approach is used to analyze and identify the essential parts in the complex biological process. The synthetic approach is used to reconstruct the biological process by putting the known parts together. Both approaches were instrumental in the development of synthetic culture media because these approaches had disadvantages relative to each other. First, the reductive approach was undertaken strenuously by Carrel and others because the synthetic approach was almost impossible without any information about the active substances for the cultivation of cells; accordingly, the substances that were important for cell culture in natural media were lined up. In contrast, the reductive approach could not be entirely successful at elucidating the complexities of the natural media because of the quantitative and qualitative limitations of the analytical method, complicated components of the natural media, and their variation from batch to batch. Accordingly, the candidate substances based on the knowledge of the reductive approach were tested by means of the synthetic approach by Eagle and others and the substances that are essential for cell culture were eventually identified. It seems to be important when and how these approaches are used for the successful development of culture media.
2.6. Identification of serum substitutes and the development of serum‐free media tailored to a cell type (1970‐)
Insulin was discovered earlier by Frederick Banting and Charles Best (1921), but full‐scale research into this peptide as a supplement for culture media began in the 1960s.63, 64 Initially, the effectiveness of insulin alone was found to be inferior to that of serum,65, 66 but the use of insulin in combination with low‐concentration serum yielded a higher level of efficacy of baby hamster kidney (BHK)21 cell growth. This finding led researchers to conclude that insulin acts in a coordinated manner with serum components. Growth factors were discovered one after another during this era: nerve growth factor,67 epidermal growth factor,68, 69 insulin‐like growth factor,70, 71, 72, 73 fibroblast growth factor (FGF),74, 75 platelet‐derived growth factor,76, 77, 78 and transforming growth factor (TGF).79, 80 The addition of these growth factors to a culture medium increased cellular proliferation. Nevertheless, their effect on cell proliferation, as with insulin, was found to be almost always inferior to the effect of serum.81, 82, 83, 84, 85
Under these circumstances, in 1976, three key reports were published that accelerated the development of serum‐free media. Ham's group discovered that a trace element of selenite is necessary for the serum‐free cultivation of human diploid cells86 and Larry J. Guilbert and Iscove showed that, besides selenite, a combination of transferrin and albumin is a good serum substitute.87 Izumi Hayashi and Gordon H. Sato discovered that a combination of several hormones and growth factors is an effective serum substitute.58 Prompted by their discoveries, attempts at serum‐free culture by using serum substitutes (eg, several hormones and growth factors, transferrin, and selenite; Table 3) grew in number and a variety of serum‐free media was developed, with each medium tailored to researchers' cell type of interest.
Table 3.
Characteristics and limitations of serum substitutes
| Category | Name | Characteristic | Limitations |
|---|---|---|---|
| Serum, tissue extracts | For example, fetal bovine serum protein,bovine pituitary extract | Contain various components, including proteins and lipids of serum or tissue origin and contribute to improved cellular proliferation and survival | Composition is undefined and therefore there is large lot‐to‐lot variation and a high risk of contamination; for example, by viruses |
| Hydrolysates | For instance, animal‐derived (animal tissues, milk), microorganism‐derived (yeast), plant‐derived (soy, wheat, rice) | Supply cells with vitamins, lipids, inorganic salts, low‐molecular‐weight peptides, and amino acids. Confirmed efficacy for culturing Chinese hamster ovary, hybridoma, baby hamster kidney, Vero, and lymph cells. Purification of antibodies and recombinant proteins is simplified because the components in question contain only low‐molecular‐weight substances, owing to ultrafiltration. Very low‐cost, as compared with serum | Composition is undefined; thus, there is a large lot‐to‐lot variation. A risk of contamination, for example, by viruses when the origin is from animals. The risk is non‐zero even when the origin is plants: for instance, when in contact with animals or animal‐source products during cultivation or the manufacturing process. Caution is necessary because the raw materials could have been exposed to high concentrations of pesticides or herbicides |
| Growth factors | For example, EGF, FGF, IGF, NGF, PDGF, TGF | Act in small quantities on cells; for example, to induce proliferation, differentiation, migration, secretion, or import. Many cells require supplementation of the medium with growth factors under serum‐free conditions. Acidic FGF readily degrades if heparin sulfate is not present on the surface of the target cells; therefore, heparin (or synthetic dextran as a substitute) is added to the medium in some situations | Animal‐derived growth factors pose a risk of contamination; for example, by viruses. The use of recombinant proteins reduces the risk of contamination; however, the risk is non‐zero because the proteins could have been produced by means of animal‐derived enzymes in the manufacturing process and for other reasons. Released from platelets, TGF‐β acts as a growth inhibitor on many epithelial cells |
| Hormones | For example, growth hormone, insulin, hydrocortisone, triiodothyronine, estrogen, androgens, progesterone, prolactin, follicle‐stimulating hormone, gastrin‐releasing peptide | Growth hormone and insulin enhance the proliferation of a variety of cells. Hydrocortisone improves the cloning efficiency of the glial cells and fibroblasts and is necessary for the maintenance of the epidermal keratinocytes and several other endothelial cell types. Triiodothyronine is necessary for MDCK epithelial cells and is used for the pulmonary epithelium. Along with hydrocortisone and prolactin, various combinations of estrogen, androgens, and progesterone are necessary for the maintenance of the mammary epithelium | Insulin is unstable at 37°C (especially in the presence of a high concentration of cysteine) and therefore must be added to a medium at a comparatively high concentration. In addition, zinc is necessary for insulin to exert its biological action and researchers ideally should use a zinc‐supplemented medium. The hydrocortisone that is present in the fetal bovine serum acts as a growth inhibitor in high‐density cultures (many cells that are closely packed; eg, glial cells, pulmonary epithelial cells). Conversely, it sometimes promotes growth in low‐density cultures |
| Carrier proteins | Albumin, transferrin, lactoferrin, and others | Albumin is used as a carrier of a variety of substances, including lipids (eg, fatty acids, cholesterol), trace elements (eg, copper, nickel), amino acids (cysteine, tryptophan), and vitamins (pyridoxal phosphate: ie, the active form of vitamin B6). As lipids cannot dissolve in an aqueous solution alone, they are more effectively supplied to cells after the formation of complexes with albumin. In addition, albumin has toxin‐neutralizing, antioxidant, and shear stress‐reducing effects. Transferrin is used as a carrier of iron. Lactoferrin can serve as a substitute for transferrin | If these agents are serum‐derived, there is a risk of contamination; for example, by viruses. Most of the serum‐derived albumin in distribution today is purified from corn by using the cold ethanol fractionation method: the products that are prepared this way contain lower proportions of other proteins. Moreover, the levels of lipids and trace elements that are bound to albumin vary from lot to lot. Sometimes, differences between lots are observed as a result: researchers should perform batch screening before using these products. Serum‐derived transferrins include compounds of porcine, bovine, and human origin. As bovine transferrin typically has low activity, researchers must in some cases work around this issue; for example, by raising the concentration |
| Lipids and related components | Cholesterol, steroids, fatty acids (eg, palmitate, stearate, oleate, linoleate), ethanolamine, choline, inositol, and others | Serve various roles: as membrane components, in nutrient storage and transport, and in signal transduction. Many established cell lines can biosynthesize the lipids that are necessary for metabolism from acetyl coenzyme A, but adding the lipids to the medium lessens the biosynthetic load. In addition, some cells lack the enzymes that are necessary for the cholesterol‐biosynthetic pathway: a source of sterols must be added to the medium in order to culture such cells. The use of lipoproteins and albumin is the physiologically closest and most effective way to solubilize proteins and deliver them to the cells | When building a culture medium under protein‐free conditions, researchers must use ethanol, surfactants (eg, Pluronic F‐68, Tween 80), or cyclodextrin to solubilize the lipids. If ethanol is used for solubilization, one must typically add a quantity of ≤1 mL/L (v/v) in view of its negative effects on cells. Caution is advised when using surfactants and cyclodextrin: they are toxic to cells at high concentrations, result in poor lipid solubility at low concentrations, and can be removed by filtration. Many commercially available lipids are animal‐derived and their performance varies |
| Transition metals | For example, Fe, Zn, Cu, Cr, I, Co, Se, Mn, Mo | These are transition elements because they readily undergo electron transfer, so they function in the active centers of enzymes and physiologically active substances inside the cell. Se, Fe, Cu, and Zn, in particular, generally are used in cell culture. Se has an antioxidant activity in the form of selenoproteins, such as glutathione peroxidase and thioredoxin reductase | Chelating agents can serve as a substitute for the Fe carrier, transferring, in cases where it must be removed from the medium. Caution is still necessary: depending on the species and concentration of the chelating agent, not only could it be ineffective as a carrier, it might also promote the production of reactive oxygen species |
| Vitamins | Fat‐soluble vitamins (A, D, E, K), water‐soluble vitamins (eg, B1, B2, B6, B12, C, folate) | Necessary for cell division and growth as precursors of various cofactors. Vitamins C and E additionally have antioxidant effects. Vitamins are present in most of the basal media, but their types and amounts (especially of the fat‐soluble vitamins) are limited in some situations; therefore, they are added according to the needs of the cell type | Vitamins A, C, D, and E are readily degraded by air oxidation. Vitamin C, moreover, reacts with trace elements and oxidatively decomposes, sometimes generating reactive oxygen species. Vitamins A, B1, B2, B12, C, and K are readily degraded by light; vitamins B1 and B5 are easily degraded by heat. Folate has poor solubility and is partially removed during filtration sterilization in some cases. Hydroxocobalamin and vitamin C interact, promoting mutual degradation |
| Polyamines | Putrescine, spermidine, spermine | Low‐molecular‐weight, basic, physiologically active amines that exist ubiquitously in cells and promote protein or nucleic‐acid synthesis. Intracellular concentrations of polyamines are regulated and maintained both by biosynthesis or decomposition inside the cell and by transport from outside the cell | Cell growth halts if the intracellular polyamine concentrations drop too low due to a disrupted balance among polyamine biosynthesis, decomposition, and transport. Moreover, apoptosis is induced if the polyamine concentration rises too much |
| Reductants | 2‐mercaptoethanol, α‐thioglycerol, reduced glutathione | Import of cystine or cysteine into cells is necessary to maintain the intracellular redox environment and the addition of reducing agents to the culture medium of cells that lack cystine transporters (eg, lymphocytes and embryonic stem cells) converts cystine into cysteine, which the cells then are able to import | Caution is necessary when adding reducing agents in the absence of albumin: this approach can damage cells |
| Protective additives, detergents | Carboxymethyl cellulose, polyvinyl pyrrolidone, Pluronic F‐68, Tween 80, and others | Reduce the shear stress generated in stirred cultures and by pipette manipulation. Pluronic F‐68 and Tween 80 also are used as solubilizers of lipophilic substances (eg, lipids, fat‐soluble vitamins) | Surfactants sometimes show cytotoxicity, depending on their concentration |
| Adhesion factors | For example, fibronectin, laminin | Promote the adhesion of anchorage‐dependent cells to vessels | Pose a risk of viral contamination if components of biological origin are used |
EGF, epidermal growth factor; FGF, fibroblast growth factor; IGF, insulin‐like growth factor; MDCK, Madin‐Darby canine kidney; NGF, nerve growth factor; PDGF, platelet‐derived growth factor; TGF, transforming growth factor.
When developing serum‐free culture media, researchers typically test combinations of individual serum substitutes. As many cells require insulin, transferrin, and selenite,88 ITS, a supplement that contains a mixture of these three substances, has become commercially available. In addition, Hiroki Murakami et al. found that ethanolamine is essential for the serum‐free cultivation of hybridomas and developed the supplement, ITES, which consists of ITS plus ethanolamine.89 Various other supplements were designed for different cell types and cultivation purposes, with the aim of the addition of various trace elements to protein‐free culture media. These include Synthetic Serum Replacement, in which various trace elements are stabilized by ethylenediaminetetraacetic acid, citric acid, and aurintricarboxylic acid;90 the B‐27 supplement that was created for use with nerve cells (it contains progesterone, putrescine, triiodothyronine, fatty acids, vitamin E, bovine serum albumin [BSA], and glutathione, in addition to ITS);91, 92 and Knockout Serum Replacement (KSR), a formulation of various active ingredients of serum (eg, BSA, amino acids, antioxidants, ITS, and trace elements).93
2.7. Improvements to basal media (1970‐)
In addition to being a source of hormones, growth factors, carrier proteins, and lipids, serum increases the levels of various low‐molecular‐weight compounds in basal media. As a result, traditional basal media from which serum is excluded were sometimes unable to adequately support cell growth.94 For example, Barnes et al. and Murakami et al. reported that the performance of DMEM, MEM, and Ham's F‐12 as basal media in serum‐free cultures is inadequate, but that the DMEM/F‐12 medium, in which Ham's F‐12 and DMEM are combined in a 1:1 ratio, shows better performance occasionally when used for certain types of cells.95, 96 The reason seems to be the large number of constituents in Ham's F‐12 and the high concentration of several nutritional constituents in DMEM: mixing the two allows each to complement the weaknesses of the other. Moreover, Murakami et al. reported that the RDF medium—a 2:1:1 mixture of RPMI 1640, DMEM, and Ham's F‐12—yields more effective cell growth of hybridomas than does the DMEM/F12 medium.97 Mixed media, however, do not always show a level of performance that is better than that of a single medium. For example, the ferrous sulfate that is contained in Ham's F‐12 is toxic to nerve cells and nerve cells proliferate more readily at a reduced osmolarity. Thus, DMEM alone is more effective than DMEM/F‐12 in this case.91 Naturally, the composition of a basal medium that is used for serum‐free culture should be optimized for each cell type. In addition, it seems that the optimization also depends on the scale of the culture and its method.98
2.8. Medical and industrial applications of animal‐cell culture technology (1978‐)
2.8.1. Culture media for the production of recombinant pharmaceuticals
Inspired by the 1982 clinical application of recombinant human insulin expressed in Escherichia coli, researchers actively proceeded to produce growth hormones, interferon α, and other substances by using E. coli or yeast as a host. With E. coli and yeast, however, it was impossible to produce proteins with glycosylation. Animal cells thus started to be used for the production of recombinant proteins, like tissue plasminogen activator, erythropoietin, interferon β, and monoclonal antibodies. The host cells that have been used in the manufacture of biopharmaceutical products include CHO cells, mouse myeloma NS0 cells, BHK cells, human embryonic kidney 293 cells, and human retinal cells. Among these, the CHO and NS0 cells have become especially popular in the field of biopharmaceutical manufacturing for the following reasons: (1) technological advances in mass‐culture methods for these two cell lines; (2) sufficient knowledge about the safety of viruses that these two cell lines contain; and (3) remarkable advances in high‐expression sublines that were derived from these two cell lines.99
In order to enhance the efficiency of the production of biopharmaceuticals, one must increase the production rate of the target protein in a culture medium that contains none or a minimal amount of ingredients of biological origin, like serum, because they significantly hamper the process of product purification. Research in this direction has been conducted to efficiently optimize the medium's composition, for example, by means of approaches that are based on the monitoring of changes in the concentration of the medium components and byproducts in the culture,100 as well as genomics‐ and proteomics‐based approaches.101 Through such efforts, as well as host cell modifications,102 the per‐cell production yield has increased nearly 10‐fold from 1986 to 2004.103 The composition of the various culture media that are used in biopharmaceutical manufacturing today has not been disclosed for commercial reasons, but the composition of a previously reported serum‐free culture medium that is used for CHO cells is detailed in Table 4 for reference.
Table 4.
Serum‐free culture media for Chinese hamster ovary cells
| Name (author[s], year) | Basal media | Supplements | Remarks |
|---|---|---|---|
|
MCDB 301 (Hamilton and Ham 1977) |
Ham's F‐12 | Trace elements (Al, Ag, Ba, Br, Cd, Co, Cr, F, Ge, I, Mn, Mo, Ni, Rb, Se, Si, Sn, Ti, V, and Zr) | A medium with 20 trace elements that are not present in Ham's F‐12 |
|
GC3
(Gasser et al. 1985) |
Modified MEM/F‐12 | Insulin, transferrin, and selenite | Developed because Chinese hamster ovary cells could not be cultured in the MCDB301 medium |
|
WCM5 (Keen and Rapson 1995) |
IMDM | Amino acids, vitamins, transition metals (Cu and Zn), ferric citrate, insulin, ethanolamine, putrescine, Pluronic F‐68, and soy peptone | Lacking high‐molecular‐weight proteins, it was developed for use with large‐scale cultures (≥8000 L). Ferric citrate is used instead of transferrin |
|
Name unspecified (Sung and Lee 2009) |
IMDM | Amino acids, ascorbate, transition metals (Cu and Zn), ferric citrate, selenite, insulin, ethanolamine, phosphatidylcholine, hydrocortisone, putrescine, pyruvate, ascorbate, Pluronic F‐68, dextran sulfate, and a hydrolysate mixture (yeast, soy, and wheat) | The combination and concentrations of the added hydrolysates were determined by using an experimental design method. It was developed to increase antibody productivity |
2.8.2. Culture media for use with pluripotent stem cells
Since the establishment of human ES cells by James A. Thomson et al. in 1998 and human iPS cells by Shinya Yamanaka et al. in 2007, the demand for these cells has increased rapidly due to their usefulness in basic and clinical studies for regenerative medicine, as well as in a variety of possible applications, such as disease modeling, drug discovery, and cytotoxicity studies. Thus, culturing these cells in a simple, low‐cost, and highly productive way became an important issue. These cells initially were cultivated on a layer of feeder cells in a serum‐ or KSR‐supplemented medium,104 but in anticipation of clinical applications, the efforts shifted to the development of culture conditions that are feeder‐free and xeno‐free (Table 5).105, 106, 107, 108, 109, 110, 111 Among these, the E8 medium that was developed by Guokai Chen et al. has become popular. With its simple composition—DMEM/F‐12 supplemented with insulin, sodium selenite, transferrin, ascorbic acid (stable form), FGF2, TGF‐β1 (or Nodal), and sodium bicarbonate—this medium allows for the long‐term growth of human iPS cells.109 Moreover, in recent years, developments have continued in the field of culture media that contain low‐molecular‐weight compounds instead of expensive growth factors (eg, FGF2 and TGF‐β1; Table 5).112
Table 5.
Serum‐free culture media for embryonic stem/induced pluripotent stem cells
| Name (author[s], year) | Basal media | Supplements | Remarks |
|---|---|---|---|
|
Knockout DMEM (Amit et al. 2000) |
DMEM | Amino acids, bFGF, 2‐mercaptoethanol, and Knockout Serum Replacement (KSR) | A medium with lower osmotic pressure than DMEM and an added serum substitute containing animal‐source components (KSR), it is for use with mouse embryonic stem cells. The cultures require feeder cells |
|
TeSR (Ludwig et al. 2006) |
DMEM/F‐12 | Vitamins, trace elements (V, Mn, Ni, Si, Sn, Mo, Cd, Cr, Ag, Al, Ba, Co, Ge, Br, I, F, Rb, Zr), selenite, LiCl, insulin, transferrin, human serum albumin (HSA), bFGF, transforming growth factor (TGF)‐β1, γ‐aminobutyric acid, pipecolic acid, glutathione, 2‐mercaptoethanol, lipids (fatty acids, cholesterol), Pluronic F‐68, and Tween 80 | A xeno‐free medium that does not require feeder cells |
|
E8 (Chen et al. 2011) |
DMEM/F‐12 | Ascorbate‐2‐phosphate, selenite, insulin, transferrin, bFGF, TGF‐β1 or NODAL, and NaHCO3 | A TeSR‐based medium. HSA (which results in large between‐lot variation) and 2‐mercaptoethanol (which negatively affects cells) were removed and supplements were refined down to the necessary minimum |
|
(Name undefined) (Hasegawa et al. 2012) |
DMEM/F‐12 | Amino acids, ascorbate, selenite, insulin, transferrin, Wnt3a, and indole derivative (ID)‐8 (DYRK inhibitor) | The expensive bFGF and TGF‐β are replaced with Wnt3a and the low‐molecular‐weight compound ID‐8. Growth is slow, compared to conventional media |
|
(Name undefined) (Hasegawa et al. 2015) |
DMEM/F‐12 | Ascorbate, selenite, insulin, transferrin, ID‐8, GSK3β inhibitor (eg, 1‐azakenpaullone), and NFAT inhibitor (eg, tacrolimus) | Wnt is replaced with a GSK3β inhibitor and NFAT inhibitor (low‐molecular‐weight compounds), it can be manufactured cheaply, and quality management is simple |
bFGF, basic fibroblast growth factor; DYRK, dual‐specificity tyrosine‐phosphorylation‐regulated kinase; GSK, glycogen synthase kinase; NFAT, nuclear factor of activated T cells.
2.8.3. Culture media for use in assisted reproductive technology
The development of in vitro fertilization, embryo culture, and embryo transfer technologies has progressed on the basis of animal experiments, primarily on rabbits and mice.113, 114, 115, 116 In 1978, the first clinical application was successful, as implemented by Patrick C. Steptoe and Robert G. Edwards.117 The medium they preferred to use for human zygote cultures was Ham's F‐10, supplemented with serum.118 It was later revealed, however, that hypoxanthine and the trace elements in Ham's F‐10 negatively affected the embryos via the production of reactive oxygen species.119, 120, 121, 122 The metabolism of a pre‐implantation embryo differs greatly from that of somatic cells; therefore, the embryo can be cultured up to the blastocyst stage by using a simple medium that consists of a balanced salt solution that is supplemented with glucose, pyruvate, lactate, and albumin.123 The concentration of glucose that was used usually for somatic cells in those days, which was as high as 5.5 mmol L−1, turned out to be unfavorable to zygote cleavage. Therefore, glucose is often added to the culture medium for use with zygotes at the low concentration of 0.2‐0.5 mmol L−1.124 The detailed composition of the embryo culture media that is used clinically has not been published to date for commercial reasons; however, according to the analysis by Morbeck et al.,125 it resembles either the G1/G2 medium that was developed by Gardner and Lane126 or the KSOMAA medium that was developed by Biggers et al.127 More information on the culture media for assisted reproductive technology is available elsewhere.124
3. Selection of a Basal Medium
Each basal medium has been designed in each case on the basis of the cell type, the origin (animal species), and the purpose of the culturing. In fact, the medium composition can differ greatly depending on such background factors. Whether supplementation with natural products is allowed is another important presupposition for the choice of a basal medium. For example, MEM (developed by Eagle) was designed under the assumption of serum supplementation and accordingly includes only the minimum necessary components (inorganic salts, sugar, essential amino acids, and water‐soluble vitamins). In contrast, Medium 199 and Ham's F‐12, intended for serum‐free culture, contain various other components. Regarding the selection of basal media, readers can be referred to the literature and suppliers' information; for instance, cell banks' sites (eg, www.atcc.org, www.phe-culturecollections.org.uk, cellbank.nibiohn.go.jp, and cell.brc.riken.jp). Table 2 shows the basic characteristics of each medium and researchers can select several candidates after comparing them experimentally. α‐MEM, DMEM, or Ham's F‐12 often are selected for adherent cell culture; the RPMI 1640 medium for suspension cell culture; and a mixed medium, like DMEM/F‐12, for serum‐free culture.
3.1. Selection of a basal medium: the roles of the medium components
3.1.1. Serum
Serum serves as a source of amino acids, proteins, vitamins, carbohydrates, lipids, hormones, growth factors, inorganic salts, trace elements, and other compounds. It also improves the pH‐buffering capacity of the medium and helps to reduce shear stress (ie, physical damage that is caused by pipette manipulation and stirring). Furthermore, serum alters the conditions at a culture substratum, allowing the adherent cells to readily proliferate there. Fetal bovine serum (FBS) is the most popular and widely applicable serum today. Other kinds of serum are used in certain situations, including calf serum (CS) and horse serum. Researchers can select an appropriate serum type on the basis of its characteristics. Fetal bovine serum generally is rich in growth factors and contains low levels of γ‐globulins (which have a cell growth‐inhibitory activity). Thus, it is suitable for cells that are difficult to coax to proliferate in culture and for the cloning of cells. In contrast, because CS has a weak growth‐promoting activity, it has been used effectively for studies of contact inhibition on the 3T3 cell line. It is also suitable for cellular differentiation studies, in which growth factors can interfere with the results.128 Lipid levels in serum rise with increasing calf age (ie, days after birth)129 and therefore CS or adult bovine serum is sometimes selected, instead of FBS, when cells with high lipid requirements are cultured. Horse serum that is harvested from adult horses via a closed system of collection has a comparatively high homogeneity between lots. Its characteristics include a low concentration of polyamine oxidase,130 which makes polyamines; the latter have a cell‐proliferative effect and are metabolically degraded less readily.
3.1.2. Alternatives to serum
When serum supplementation is inappropriate or undesirable, researchers can choose several substitutes for serum. They include serum extracts, tissue extracts or hydrolysates, growth factors, hormones, carrier proteins like albumin and transferrin, lipids, metals, vitamins, polyamines, and reductants (Table 3). The number of combinations of these supplements is nearly infinite and they often interact with each other. Their selection thus incurs an enormous effort, time, and cost: one cannot design an optimal medium by simply trying them in arbitrary combinations at random. Thorough studies on past records of successful combinations, if available, are helpful. The following is an example of a protocol when researchers try to implement a serum‐free culture by themselves:131
Try a medium where the ITS supplement is added to DMEM/F‐12.
If using glutamine, set its concentration to 2–4 mmol L−1 and consider the stable form: L‐alanyl‐L‐glutamine (see the next section).
Add growth factors, hormones, vitamins, trace elements, and lipids according to the requirements of the cell type under study.
Pay attention to the osmotic pressure.
Use antibiotics as little as possible.
When culturing adherent cells, consider using substrates, such as fibronectin and laminin, for cell attachment.
For a stirred culture, consider supplementing the medium with protective agents like Pluronic F‐68 to minimize the shear force.
Adapt the cells to the new medium with great caution.
Verify that the cell performance has not changed in the new medium.
The serum‐free and protein‐free media that are sold by a variety of manufacturers largely show good performance and researchers can use them to culture their cells of interest. Nonetheless, their composition almost has never been disclosed for commercial reasons, and therefore, technically, such media cannot be termed “chemically defined.”
4. Points of Caution When Using Culture Media
4.1. Setting up a serum‐free culture
There are two points to note when working with a serum‐free culture system. First, unintended clones of cells in a culture vessel can be selected during subculturing because serum‐free media can promote the proliferation of a particular cell clone or subtype of cell more often than can serum‐containing media. Second, impurities in a serum‐free medium, both avoidable and inevitable, might have stronger effects on the cultured cells than can serum‐containing media because of a lack of the toxin‐neutralizing activity that serum contains. In addition, there are other precautions for serum‐free cultures: that is, the minimization of the concentration of trypsin and the careful selection of adhesion factors.57
4.2. pH changes
The pH of the culture medium that is used for animal‐cell cultures generally is maintained by the equilibrium relationship between sodium bicarbonate (NaHCO3) in the culture medium and CO2 in the incubator (Fig. 1). Many laboratories set the CO2 concentration in the incubator to 5%‐6% by convention, but technically, it should be adjusted according to the concentration of the NaHCO3 in the culture medium. For example, 5% CO2 is appropriate for a medium with 26 mmol L−1 NaHCO3 added (eg, MEM medium), 2% CO2 for a medium with 14 mmol L−1 added (eg, Ham's F‐12 medium), and 10% CO2 for a medium with 44 mmol L−1 NaHCO3 added (eg, DMEM medium). These numbers are theoretical values that are calculated by using the Henderson–Hasselbalch equation (Fig. 1): in practice, checking the pH of a culture medium after it reaches equilibrium and then making minute adjustments to the CO2 concentration are preferable.
Figure 1.

The pH control mechanism of culture media, based on the bicarbonate buffer system and the Henderson–Hasselbalch equation. When dissolved in water, sodium bicarbonate (NaHCO 3) dissociates to form a sodium ion (Na+) and a bicarbonate ion (HCO 3 −). The latter reacts with H+ in solution to form carbonic acid (H2 CO 3), which dissociates into CO 2 and H2O. These two reactions attain their respective equilibria. The CO 2 in solution also reaches equilibrium with CO 2 in the gas phase. As a result, increasing the concentration of gas phase CO 2 increases the amount of CO 2 that is dissolved in the culture medium, in turn raising the H2 CO 3 concentration and lowering the pH. In contrast, if the concentration of the gas phase CO 2 is lowered, then the pH rises due to the reverse reaction. The relationship between the culture medium pH and the concentrations of CO 2 and NaHCO 3 can be expressed by the Henderson–Hasselbalch equation: pH=pKa+log[HCO 3 −]/[CO 2]Liquid phase, where: pKa is the negative log of the acid dissociation constant.
The 4‐(2‐hydroxyethyl)‐1‐piperazineethanesulfonic acid (HEPES), with its strong buffering capacity, might have buffering effects in a medium. Especially when cultured cells are handled outside the CO2 incubator, the pH of the medium with HEPES is more stable than with only bicarbonate. It also might be beneficial for cultures of high cell density; for instance, when the pH can drop rapidly due to an accumulation of metabolites, such as lactate. With serum‐free media, which lack the buffering capacity of serum, the pH‐buffering role of HEPES could be more relevant. Nevertheless, HEPES might have negative effects on certain types of cells, such as chick embryo epiphyseal chondrocytes and ES cells.107, 132 It also has been demonstrated that HEPES‐containing media produce an increased level of cytotoxic products, mostly hydrogen peroxide, during exposure of the medium to visible light.133 Researchers should be aware of such drawbacks of HEPES and confirm that the addition of HEPES is harmless to the cells that they use.
Phenol red, added to a medium as a pH indicator, sometimes affects the proliferation of cells and shows estrogenic activity.134, 135 Therefore, investigators should not forget to test whether such characteristics of phenol red affect their cells and empirical results, especially when estrogen‐sensitive cells, like mammary gland cells, are involved.
4.3. Oxidative stress
Culture experiments in general and especially when performed under a higher oxygen concentration, such as organ culture experiments, can expose cells to oxidative stress, which negatively affects the cells and tissues in culture. Supplementation with substances with an antioxidant activity (eg, vitamins [C, E], glutathione, selenite, β‐mercaptoethanol, dithiothreitol, or lipoic acid) is effective, especially when serum (it contains antioxidants) is not added to the medium. In addition, iron and copper ions in a free state promote the production of reactive oxygen species. It is therefore best for these ions to be complexed with appropriate carriers (eg, transferrin, albumin, or chelating agents) and to be supplied to the cells in this state, while they are isolated from reactive systems. Caution is also necessary with respect to free iron ions, which are readily hydroxylated and precipitate in an aqueous solution.
4.4. Nutrient requirements
Sufficient amounts of nutrients in the medium are a prerequisite for cells to behave properly. Some cell types require higher levels of nutrients than others do, depending on their metabolic activity and proliferation rate. Such characteristics of cells should be taken into account in the selection of a medium. DMEM, for example, was designed originally to contain glucose at 5.6 mmol L−1. Now, a modified version of DMEM with a high glucose concentration, 25 mmol L−1, which can be used for cells requiring greater amounts of glucose, is available from various suppliers. One caveat when using this high‐glucose medium for actively proliferating cells is the accumulation of metabolites, like lactate, and a plunge in the pH. Researchers are advised to replace the medium at proper intervals or to use HEPES to confer a stronger pH‐buffering capacity onto the medium.
Glutamine, one of the essential amino acids, is an energy source for mammalian cells in culture, in addition to being a biosynthetic material for nucleic acids and proteins. The glutamine requirements for cells in culture are ~3‐40‐fold greater than those of other amino acids.40 During the cultivation of cells with high nutrient requirements, the addition of glutamine could be helpful. In contrast, glutamine readily decomposes in culture media and generates cytotoxic ammonia. Consequently, researchers may consider adding glutamine to the medium immediately before use or to consider using glutamine derivatives, such as L‐alanyl‐L‐glutamine or glycyl‐L‐glutamine, which are resistant to degradation.
Non‐essential amino acids (NEAAs) can be biosynthesized in the cell. Thus, NEAAs are not included in some basal media, such as BME and MEM. Nevertheless, not every cell type can produce sufficient amounts of NEAAs in the cultured state. The addition of NEAAs to the medium, therefore, could ensure more favorable culture conditions. Even cells that can biosynthesize NEAAs at a necessary rate might benefit from the addition of NEAAs to the medium because this action alleviates the biosynthetic load of the cells.
Branched‐chain amino acids (BCAAs), including valine, leucine, and isoleucine, require special attention. They all belong to the group of essential amino acids and several cell types, including human fibroblasts and mouse myeloma cells, require larger amounts of BCAAs. For those cells, increased concentrations of BCAAs or replacing the medium at proper intervals can be considered in order to obtain the best results of an experiment.
5. Current Issues with Culture Media
5.1. Design of experiments for the optimization of the medium components
Most of the culture media that generally are used today consist of a basal medium and several supplements. Researchers pay no attention to the possible interactions among the components of the basal medium and these supplements. The fact is that each component of a culture medium does not always act in isolation. The components can interact and their effects on the cells should be expected to be influenced by those interactions. Additionally, the concentration of a medium component and its effect on the cells might not follow a linear dependence. Cells' response to each component could be curvilinear, as shown in Figure 2. Therefore, the best way to find the optimal concentration and interactions of each component in a culture medium might be a statistical experimental strategy called a “design of experiments” (DoE), rather than the classical one‐factor‐at‐a‐time experiments. The latter approach might miss the optimal point of concentration because there are some unexamined areas in the range of experimental settings (Fig. 3A). By contrast, the DoE can allocate design points, namely the concentrations of the components to be examined, evenly throughout the area of settings. Then, the obtained empirical data will be applied to the appropriate mathematical models, such as the general linear model or the logistic regression model. In general, when the data corresponding to the response variable (also called the “objective variable”) are continuous, such as the production volume of a biopharmaceutical, the general linear model is used. With binominal data, such as the survival rate of cells, the logistic regression model is preferable. By means of these models, an optimal concentration of each component (which is expected to induce the best response of the cultured cells) will be estimated (Fig. 3B). The DoE mainly consists of screening designs and response surface designs. A screening design is used first when one is trying to extract the most important components from many, say more than five, candidates of the initial components. The strength of the screening designs, such as fractional factorial or the Plackett–Burman design, is that they can reduce the number of experimental runs significantly, in comparison with the number that otherwise has to be done with the full factorial design, which involves every possible combination of components. For example, when trying to find the key factors in Medium 199, which consists of 60 components, only 64 experiments are needed with the Plackett–Burman design, whereas 260 experiments are required with the full factorial design. Once the key factors are identified, preferably eight or fewer, a response surface design, such as the central composite design or Box–Behnken design, will be used to estimate a non‐linear response of the cells and to identify the optimized concentrations and interactions of each component. These approaches have become a powerful tool for the improvement of productivity in the field of cell culture.136 Moreover, a new screening design, called a “definitive screening design,” recently was reported, with the claim that it has the advantages of both a screening design and a response surface design.137 This design enables the estimation of not only the main effects (the effect of a certain component alone) but also the interactions of the components and the factors with non‐linear effects, at a minimal number of experimental runs. In the case of Medium 199, only 121 experiments are needed. This approach could be especially useful for studies where the sample size is limited, such as a primary culture, organ culture, and embryo culture, although there are some restrictions in this design. The establishment of effective development systems for culture media that combine in vitro and in silico approaches and the optimization of the combinations of all the components in the culture medium are crucial for further improvements in cell culture performance.
Figure 2.
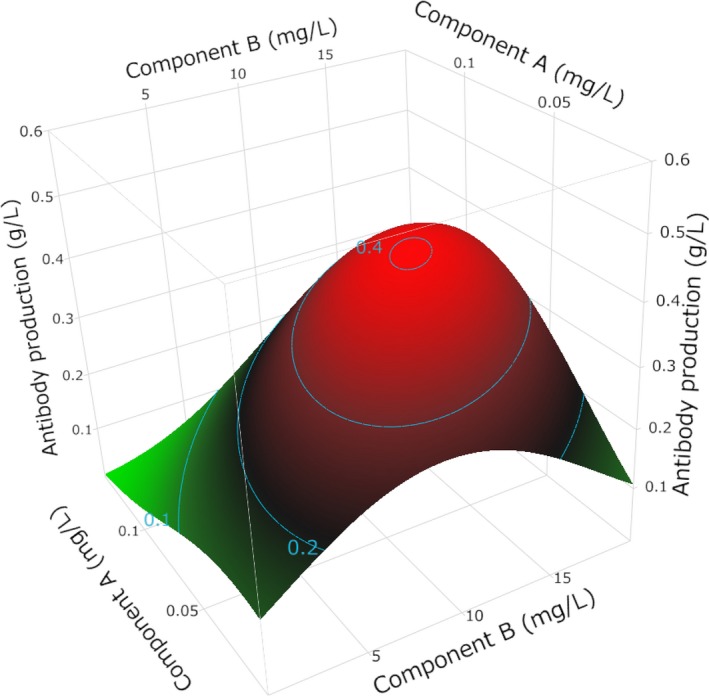
An example of a concentration–response surface. When Component B's concentration is low (eg, 0mg/L), antibody production falls as Component A's concentration rises; however, when Component B's concentration is high (eg, 20mg/L), antibody production increases as Component A's concentration rises. Such a phenomenon—one component influencing the response of another component—is called a “two‐factor interaction.” The relationship between the concentration and response is not necessarily linear, as is the case for Component B. An analysis of the concentration by using at least a three‐level screening design is necessary to understand such responses
Figure 3.

Concepts of the one‐factor‐at‐a‐time experiment and design of experiment (DoE). These figures show the difference in strategies between a one‐factor‐at‐a‐time experiment and a DoE for the same experimental runs. A, In the case of the one‐factor‐at‐a‐time experiment, the optimal concentration of one component (eg, component A) is determined at a fixed concentration of another component (eg, component B). Then, the optimal concentration of component B is determined at the optimal concentration of component A. This strategy, which usually has been used, has a big disadvantage of missing the optimal point because there are some unexamined areas in the range of parameters. B, In contrast, the DoE is a model‐based statistical method that can clarify the relationship between the response of the cells and the concentrations of the tested components in the range of settings. The process of the DoE is mainly composed of four steps. First, allocate the design points evenly throughout the area. Second, record the response of the cells for each run. Third, fit the collected data to an appropriate model (eg, a logistic regression model for a binomial response) and validate the relevance of the model to decide whether it is available for the next step. Finally, use the model to optimize the concentrations of the components or to predict a response of the cells
5.2. Undefined medium supplements
Supplements of biogenic origin, like serum, can be a cause of variation in the experimental results from batch to batch. They also carry a risk of microbial contamination of the culture medium. Thus, the replacement of those supplements with defined ones has been pursued in the history of culture media, as described above.43, 87 At the dawn of the technology, a human embryo was cultured in a medium containing serum. In the mid‐1980s, the serum could be replaced by serum albumin.138 Albumin is the most abundant protein in serum and is multifunctional. It binds to various water‐insoluble substances like lipids. The lipids that are carried by albumin become an energy source and biosynthetic substances for an embryo.139, 140, 141 In addition, serum albumin serves as an antioxidant, osmotic regulator, and neutralizer of toxins. These functions are key benefits that serum usually provides to the media. Nonetheless, the use of serum albumin in place of serum has not contributed much to the development of chemically defined media. First, most, if not all, commercial serum albumin versions contain >100 serum proteins, although these admixtures have very low concentrations.142, 143 Second, albumin can bind to potential toxins, like phthalates144 (common plasticizers) or endotoxins. Thus, those toxins can be present in the commercial versions of serum albumin. These impurities in serum albumin products even were reported to vary in concentration from batch to batch, thus affecting the results of the embryo culture.145, 146, 147, 148, 149, 150 Therefore, the development of the culture media without undefined supplements is desirable, especially for human embryos. Recently, highly purified recombinant human albumin became commercially available. It is worth trying it as a substitute for serum‐derived albumin. Summarized in Table 3 are the points of caution when researchers use undefined supplements.
5.3. Contamination of a medium
Foreign substances from unidentified sources can contaminate a culture medium, thus possibly affecting the empirical results. Typically, those contaminants include viruses, bacteria, mycoplasma, and endotoxins. There are, however, other types of contaminants, like plasticizers that might be eluted from plastic instruments144 or trace elements, even in water. These substances also can affect the cells in culture.60 It also was reported that some toxic substances are eluted from the microfilters that are used for sterilization.151 Some of this contamination seems to be even inevitable, but care must be taken to minimize it in order to make culture experiments reliable and highly reproducible. Thus, researchers may consider practicing the sterile technique strictly and selecting culture instruments carefully. Washing the instruments with the culture medium immediately before use is recommended in some special cases.151, 152, 153
6. Conclusion
Ever since Harrison's successful cultivation of animal cells, cell culture technology has developed in leaps and bounds, with many breakthroughs. With the consumables (eg, culture media) and cell culture equipment now supplied on a commercial basis, it became possible for anyone to work easily with cultured cells. As a result, there have been fewer opportunities lately to appreciate the research value of culture media, as well as their shortcomings and limitations. With progress in regenerative medicine and biopharmaceuticals, the creation of culture systems that do not require a human intervention is expected to continue: this trend will probably intensify in the future. Even with this trend, the culture medium is of paramount importance for the best quality of cell culture experiment, as well as biopharmaceutical work. Here again, it should be noted that the current culture media and their formulations have been established through the timeless efforts of innumerable researchers. From now on, investigators should drive the further evolution of culture media, with the aim of improving culture performance.
Disclosures
Conflict of interest: The authors declare no conflict of interest. Human and Animal Rights: This article does not contain any study on human or animal participants that was performed by any of the authors.
Acknowledgements
We are indebted to Akio Matsuhisa for critical reading of the manuscript. We are especially thankful to Takehiko Ogawa for his advice and expertise.
Yao T, Asayama Y. Animal‐cell culture media: History, characteristics, and current issues. Reprod Med Biol. 2017;16:99–117. https://doi.org/10.1002/rmb2.12024
References
- 1. Keehan WM, Barnes D, Reid L, Stanbridge E, Murakami H, Sato G. Frontiers in mammalian cells culture. In Vitro Cell Dev Biol. 1990;26:9–23. [DOI] [PubMed] [Google Scholar]
- 2. Brunner D, Frank J, Appl H, Schoffl H, Pfaller W, Gstraunthaler G. Serum‐free cell culture: the serum‐free media interactive online database. ALTEX. 2010;27:53–62. [DOI] [PubMed] [Google Scholar]
- 3. Freshney RI. Serum‐free media In: Freshney RI, ed. Culture of Animal Cells. Hoboken, NJ: John Wiley & Sons, Inc.; 2010:115–132. [Google Scholar]
- 4. Ringer S. Concerning the influence exerted by each of the constituents of the blood on the contraction of the ventricle. J Physiol. 1882;3:380–393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Ringer S. A further contribution regarding the influence of the different constituents of the blood on the contraction of the heart. J Physiol. 1883;4:29–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Locke FS, Rosenheim O. Contributions to the physiology of the isolated heart: the consumption of dextrose by mammalian cardiac muscle. J Physiol. 1907;36:205–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Tyrode M. The mode of action of some purgative salts. Arch Int Pharmacodyn. 1910;20:205–223. [Google Scholar]
- 8. Krebs H, Henseleit K. Untersuchungen über die harnstoffbildung im tierkörper. Klin Wochenschr. 1932;11:757–759. [Google Scholar]
- 9. Gey GO, Gey MK. The maintenance of human normal cells and tumor cells in continuous culture: I. Preliminary report: cultivation of mesoblastic tumors and normal tissue and notes on methods of cultivation. Am J Cancer. 1936;27:45–76. [Google Scholar]
- 10. Earle W, Schilling E, Stark T, Straus N, Brown M, Shelton E. Production of malignancy in vitro; IV: the mouse fibroblast cultures and changes seen in the living cells. J Natl Cancer Inst. 1943;4:165–212. [Google Scholar]
- 11. Hanks JH, Wallace RE. Relation of oxygen and temperature in the preservation of tissues by refrigeration. Proc Soc Exp Biol Med. 1949;71:196–200. [DOI] [PubMed] [Google Scholar]
- 12. Roux W. Beitriige zur entwicklungsmechanik des embryo. Z Biol. 1885;21:411. [Google Scholar]
- 13. Loeb L. Über die enstehung von bindegewebe, leucocyten und roten blutkörperchen aus epithel und über eine methode In: Loeb L, ed. Isolierte Gewebsteile zu Züchten. Chicago: Druck von Max Stern & Company; 1897:1–56. [Google Scholar]
- 14. Jolly J. Sur la durée de la vie et de la multiplication des cellules animales en dehors de l'organisme. Compt Rend Soc Biol Paris. 1903;55:1266–1268. [Google Scholar]
- 15. Harrison RG, Greenman MJ, Mall FP, Jackson CM. Observations of the living developing nerve fiber. Anat Rec. 1907;1:116–128. [Google Scholar]
- 16. Carrel A. A method for the physiological study of tissues in vitro. J Exp Med. 1923;38:407–418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Burrows MT. The cultivation of tissues of the chick‐embryo outside the body. JAMA. 1910;55:2057–2058. [Google Scholar]
- 18. Carrel A, Burrows MT. Cultivation of tissues in vitro and its technique. J Exp Med. 1911;13:387–396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Carrel A. On the permanent life of tissues outside of the organism. J Exp Med. 1912;15:516–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Carrel A. Artificial activation of the growth in vitro of connective tissue. J Exp Med. 1913;17:14–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Ebeling AH. A ten year old strain of fibroblasts. J Exp Med. 1922;35:755–759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Corner GW. Warren Harmon Lewis. Biograph. Memoirs Natl Acad. Sci. 1967;39:323–358. [Google Scholar]
- 23. Lewis MR, Lewis WH. The cultivation of tissues from chick embryos in solutions of NaCl, CaCl2, KCl and NaHCO3 . Anat Rec. 1911;5:277–293. [Google Scholar]
- 24. Lewis WH, Lewis MR. The cultivation of chick tissues in media of known chemical constitution. Anat Rec. 1912;6:207–211. [Google Scholar]
- 25. Lewis MR. The importance of dextrose in the medium of tissue cultures. J Exp Med. 1922;35:317–322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Baker LE, Carrel A. Action on fibroblasts of the protein fraction of embryonic tissue extract. J Exp Med. 1926;44:387–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Carrel A, Baker LE. The chemical nature of substances required for cell multiplication. J Exp Med. 1926;44:503–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Baker LE, Carrel A. Effect of the amino acids and dialyzable constituents of embryonic tissue juice on the growth of fibroblasts. J Exp Med. 1926;44:397–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Baker LE. The chemical nature of the substances required for cell multiplication: II. Action of glutathione, hemoglobin, and ash of liver on the growth of fibroblasts. J Exp Med. 1929;49:163–182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Vogelaar JPM, Erlichman E. A feeding solution for cultures of human fibroblasts. Am J Cancer. 1933;18:28–38. [Google Scholar]
- 31. Baker LE. Artificial media for the cultivation of fibroblasts, epithelial cells and monocytes. Science. 1936;83:605–606. [DOI] [PubMed] [Google Scholar]
- 32. Hayflick L, Moorhead PS. The serial cultivation of human diploid cell strains. Exp Cell Res. 1961;25:585–621. [DOI] [PubMed] [Google Scholar]
- 33. Sanford KK, Earle WR, Likely GD. The growth in vitro of single isolated tissue cells. J Natl Cancer Inst. 1948;9:229–246. [PubMed] [Google Scholar]
- 34. Gey G, Coffman W, Kubicek M. Tissue culture studies of the proliferative capacity of cervical carcinoma and normal epithelium. Cancer Res. 1952;12:264–265. [Google Scholar]
- 35. Fischer A, Astrup T, Ehrensvard G, Oehlenschlager V. Growth of animal tissue cells in artificial media. Proc Soc Exp Biol Med. 1948;67:40–46. [DOI] [PubMed] [Google Scholar]
- 36. Fischer A. Amino‐acid metabolism of tissue cells in vitro. Biochem J. 1948;43:491–497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Eagle H. The specific amino acid requirements of a mammalian cell (strain L) in tissue culture. J Biol Chem. 1955;214:839–852. [PubMed] [Google Scholar]
- 38. Eagle H. The specific amino acid requirements of a human carcinoma cell (strain HeLa) in tissue culture. J Exp Med. 1955;102:37–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Eagle H. Nutrition needs of mammalian cells in tissue culture. Science. 1955;122:501–514. [DOI] [PubMed] [Google Scholar]
- 40. Eagle H. Amino acid metabolism in mammalian cell cultures. Science. 1959;130:432–437. [DOI] [PubMed] [Google Scholar]
- 41. Dulbecco R, Freeman G. Plaque production by the polyoma virus. Virology. 1959;8:396–397. [DOI] [PubMed] [Google Scholar]
- 42. Stanners CP, Eliceiri GL, Green H. Two types of ribosome in mouse–hamster hybrid cells. Nature New Biol. 1971;230:52–54. [DOI] [PubMed] [Google Scholar]
- 43. Iscove NN, Melchers F. Complete replacement of serum by albumin, transferrin, and soybean lipid in cultures of lipopolysaccharide‐reactive b lymphocytes. J Exp Med. 1978;147:923–933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Neuman RE, McCoy TA. Growth‐promoting properties of pyruvate, oxaloacetate, and α‐ketoglutarate for isolated Walker carcinosarcoma 256 cells. Exp Biol Med. 1958;98:303–306. [DOI] [PubMed] [Google Scholar]
- 45. McCoy TA, Maxwell M, Kruse PF. The amino acid requirements of the Jensen sarcoma in vitro. Cancer Res. 1959;19:591–595. [PubMed] [Google Scholar]
- 46. Moore GE, Ito E, Ulrich K, Sandberg AA. Culture of human leukemia cells. Cancer. 1966;19:713–723. [DOI] [PubMed] [Google Scholar]
- 47. Moore GE, Gerner RE, Franklin HA. Culture of normal human leukocytes. JAMA. 1967;199:519–524. [PubMed] [Google Scholar]
- 48. White PR. Cultivation of animal tissues in vitro in nutrients of precisely known constitution. Growth. 1946;10:231–289. [PubMed] [Google Scholar]
- 49. Jacoby F, Darke SJ. Animal tissue culture with a synthetic medium. Nature. 1948;161:768–769. [DOI] [PubMed] [Google Scholar]
- 50. Morgan JF, Morton HJ, Parker RC. Nutrition of animal cells in tissue culture. I. Initial studies on a synthetic medium. Exp Biol Med. 1950;73:1–8. [DOI] [PubMed] [Google Scholar]
- 51. Parker RC, Castor LN, McCulloch EA. Altered cell strains in continuous culture. A general survey. Spec Publ NY Acad Sci. 1957;5:303–313. [Google Scholar]
- 52. Parker RC. Chemically defined media In: Parker RC, ed. Methods of Tissue Culture. New York: Harper & Row; 1961:62–80. [Google Scholar]
- 53. Evans VJ, Bryant JC, McQuilkin WT, et al. Studies of nutrient media for tissue cells in vitro II. An improved protein‐free chemically defined medium for long‐term cultivation of strain L‐929 cells. Cancer Res. 1956;16:87–94. [PubMed] [Google Scholar]
- 54. Waymouth C. Rapid proliferation of sublines of nctc clone 929 (strain L) mouse cells in a simple chemically defined medium (MB 752/1). J Natl Cancer Inst. 1959;22:1003–1017. [DOI] [PubMed] [Google Scholar]
- 55. Ham RG. Albumin replacement by fatty acids in clonal growth of mammalian cells. Science. 1963;140:802–803. [DOI] [PubMed] [Google Scholar]
- 56. Ham RG. Putrescine and related amines as growth factors for a mammalian cell line. Biochem Biophys Res Commun. 1963;14:34–38. [DOI] [PubMed] [Google Scholar]
- 57. Ham RG. Clonal growth of mammalian cells in a chemically defined, synthetic medium. Proc Natl Acad Sci USA. 1965;53:288–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Hayashi I, Sato GH. Replacement of serum by hormones permits growth of cells in a defined medium. Nature. 1976;259:132–134. [DOI] [PubMed] [Google Scholar]
- 59. Murakami H. Serum‐free media used for cultivation of hybridomas. Adv Biotechnol Processes. 1989;11:107–141. [PubMed] [Google Scholar]
- 60. Hamilton WG, Ham RG. Clonal growth of Chinese hamster cell lines in protein‐free media. In Vitro Cell Dev Biol. 1977;13:537–547. [DOI] [PubMed] [Google Scholar]
- 61. Takaoka T, Katsuta H. Long‐term cultivation of mammalian cell strains in protein‐ and lipid‐free chemically defined synthetic media. Exp Cell Res. 1971;67:295–304. [DOI] [PubMed] [Google Scholar]
- 62. Oshima M, Miyamoto I, Takaoka T, Goto K, Kagawa Y. Unsaturated fatty acids in mammalian cell strains cultured for more than 10 years in serum‐free synthetic media. Jpn J Exp Med. 1987;57:351–354. [PubMed] [Google Scholar]
- 63. Lieberman I, Ove P. Growth factors for mammalian cells in culture. J Biol Chem. 1959;234:2754–2758. [PubMed] [Google Scholar]
- 64. Higuchi K. Studies on the nutrition and metabolism of animal cells in serum‐free media: I. Serum‐free monolayer cultures. J Infect Dis. 1963;112:213–220. [DOI] [PubMed] [Google Scholar]
- 65. Clarke GD, Stoker MGP, Ludlow A, Thornton M. Requirement of serum for DNA synthesis in BHK 21 cells: effects of density, suspension and virus transformation. Nature. 1970;227:798–801. [DOI] [PubMed] [Google Scholar]
- 66. Clarke GD, Stoker MGP. Conditions affecting the response of cultured cells to serum In: Wolstenholme GEW, Knight J, eds. Ciba Foundation Symposium – Growth Control in Cell Cultures. Chichester: John Wiley & Sons, Ltd.; 1971:17–32. [Google Scholar]
- 67. Levi‐Montalcini R. Effects of mouse tumor transplantation on the nervous system. Ann NY Acad Sci. 1952;55:330–344. [DOI] [PubMed] [Google Scholar]
- 68. Cohen S. Isolation of a mouse submaxillary gland protein accelerating incisor eruption and eyelid opening in the new‐born animal. J Biol Chem. 1962;237:1555–1562. [PubMed] [Google Scholar]
- 69. Taylor JM, Mitchell WM, Cohen S. Epidermal growth factor: physical and chemical properties. J Biol Chem. 1972;247:5928–5934. [PubMed] [Google Scholar]
- 70. Salmon WD Jr, Daughaday WH. A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro. J Lab Clin Med. 1957;49:825–836. [PubMed] [Google Scholar]
- 71. Froesch ER, Buergi H, Ramseier EB, Bally P, Labhart A. Antibody‐suppressible and nonsuppressible insulin‐like activities in human serum and their physiologic significance. An insulin assay with adipose tissue of increased precision and specificity. J Clin Invest. 1963;42:1816–1834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Daughaday WH, Hall K, Raben MS, Salmon WD Jr, van den Brande JL, van Wyk JJ. Somatomedin: proposed designation for sulphation factor. Nature. 1972;235:107. [DOI] [PubMed] [Google Scholar]
- 73. Rinderknecht E, Humbel RE. Polypeptides with nonsuppressible insulin‐like and cell‐growth promoting activities in human serum: Isolation, chemical characterization, and some biological properties of forms I and II. Proc Natl Acad Sci USA. 1976;73:2365–2369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Gospodarowicz D. Localisation of a fibroblast growth factor and its effect alone and with hydrocortisone on 3t3 cell growth. Nature. 1974;249:123–127. [DOI] [PubMed] [Google Scholar]
- 75. Gospodarowicz D. Purification of a fibroblast growth factor from bovine pituitary. J Biol Chem. 1975;250:2515–2520. [PubMed] [Google Scholar]
- 76. Kohler N, Lipton A. Platelets as a source of fibroblast growth‐promoting activity. Exp Cell Res. 1974;87:297–301. [DOI] [PubMed] [Google Scholar]
- 77. Ross R, Glomset J, Kariya B, Harker L. A platelet‐dependent serum factor that stimulates the proliferation of arterial smooth muscle cells in vitro. Proc Natl Acad Sci USA. 1974;71:1207–1210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Antoniades HN, Scher CD, Stiles CD. Purification of human platelet‐derived growth factor. Proc Natl Acad Sci USA. 1979;76:1809–1813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. De Larco JE, Preston YA, Todaro GJ. Properties of a sarcoma‐growth‐factor‐like peptide from cells transformed by a temperature‐sensitive sarcoma virus. J Cell Physiol. 1981;109:143–152. [DOI] [PubMed] [Google Scholar]
- 80. Anzano MA, Roberts AB, Meyers CA, et al. Communication: synergistic interaction of two classes of transforming growth factors from murine sarcoma cells. Cancer Res. 1982;42:4776–4778. [PubMed] [Google Scholar]
- 81. Hollenberg MD, Cuatrecasas P. Epidermal growth factor: receptors in human fibroblasts and modulation of action by cholera toxin. Proc Natl Acad Sci USA. 1973;70:2964–2968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Gospodarowicz D, Moran JS. Stimulation of division of sparse and confluent 3t3 cell populations by a fibroblast growth factor, dexamethasone, and insulin. Proc Natl Acad Sci USA. 1974;71:4584–4588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Holley RW, Kiernan JA. Control of the initiation of DNA synthesis in 3t3 cells: serum factors. Proc Natl Acad Sci USA. 1974;71:2908–2911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Smith GL, Temin HM. Purified multiplication‐stimulating activity from rat liver cell conditioned medium: comparison of biological activities with calf serum, insulin, and somatomedin. J Cell Physiol. 1974;84:181–192. [DOI] [PubMed] [Google Scholar]
- 85. Gospodarowicz D, Moran J. Optimal conditions for the study of growth control in balb/c 3t3 fibroblasts. Exp Cell Res. 1975;90:279–284. [DOI] [PubMed] [Google Scholar]
- 86. McKeehan WL, Hamilton WG, Ham RG. Selenium is an essential trace nutrient for growth of wi‐38 diploid human fibroblasts. Proc Natl Acad Sci USA. 1976;73:2023–2027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Guilbert LJ, Iscove NN. Partial replacement of serum by selenite, transferrin, albumin and lecithin in haemopoitec cell cultures. Nature. 1976;263:594–595. [DOI] [PubMed] [Google Scholar]
- 88. Barnes D, Sato G. Serum‐free cell culture: a unifying approach. Cell. 1980;22:649–655. [DOI] [PubMed] [Google Scholar]
- 89. Murakami H, Masui H, Sato GH, Sueoka N, Chow TP, Kano‐Sueoka T. Growth of hybridoma cells in serum‐free medium: ethanolamine is an essential component. Proc Natl Acad Sci USA. 1982;79:1158–1162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Bertheussen K. Growth of cells in a new defined protein‐free medium. Cytotechnology. 1993;11:219–231. [DOI] [PubMed] [Google Scholar]
- 91. Brewer GJ, Torricelli JR, Evege EK, Price PJ. Optimized survival of hippocampal neurons in b27‐supplemented neurobasal™, a new serum‐free medium combination. J Neurosci Res. 1993;35:567–576. [DOI] [PubMed] [Google Scholar]
- 92. Brewer GJ. Serum‐free b27/neurobasal medium supports differentiated growth of neurons from the striatum, substantia nigra, septum, cerebral cortex, cerebellum, and dentate gyrus. J Neurosci Res. 1995;42:674–683. [DOI] [PubMed] [Google Scholar]
- 93. Price PJ, Goldsborough MD, Tilkins ML. Embryonic stem cell serum replacement. International Patent Application, 1998, WO 98/30679.
- 94. Keen MJ, Rapson NT. Development of a serum‐free culture medium for the large scale production of recombinant protein from a Chinese hamster ovary cell line. Cytotechnology. 1995;17:153–163. [DOI] [PubMed] [Google Scholar]
- 95. Barnes D, Sato G. Growth of a human mammary tumour cell line in a serum‐free medium. Nature. 1979;281:388–389. [DOI] [PubMed] [Google Scholar]
- 96. Murakami H, Edamoto T, Nakamura H, Omura H. Growth of myeloma mpc‐11 cells in serum‐free growth factor supplemented medium. Agric Biol Chem. 1982;46:1831–1837. [Google Scholar]
- 97. Murakami H. Serum‐free cultivation of plasmacytomas and hybridomas In: Barnes D, Sirbasku D, Sato G, eds. Methods for Serum‐Free Culture of Neuronal and Lymphoid Cells. New York: Alan R. Liss, Inc.; 1984:197–206. [Google Scholar]
- 98. Murakami H, Yamada K. Production of cancer specific monoclonal antibodies with human–human hybridomas and their serum‐free, high density, perfusion culture In: Spier R, Griffiths J, eds. Modern Approaches to Animal Cell Technology. London: Butterworths; 1987:52–76. [Google Scholar]
- 99. Zhu MM, Mollet M, Hubert RS. Industrial production of therapeutic proteins: cell lines, cell culture, and purification In: Kent AJ, ed. Handbook of Industrial Chemistry and Biotechnology. Boston: Springer; 2012:1229–1248. [Google Scholar]
- 100. Lu F, Toh PC, Burnett I, et al. Automated dynamic fed‐batch process and media optimization for high productivity cell culture process development. Biotechnol Bioeng. 2013;110:191–205. [DOI] [PubMed] [Google Scholar]
- 101. Aboytes KA, Allison DW, Donahue LM, Fong D, Johnson TK. Genomic and proteomic approaches for the development of cell culture medium. International Patent Application, 2004, WO 2004/101808 A2.
- 102. Kim JY, Kim Y‐G, Lee GM. CHO cells in biotechnology for production of recombinant proteins: current state and further potential. Appl Microbiol Biotechnol. 2012;93:917–930. [DOI] [PubMed] [Google Scholar]
- 103. Wurm FM. Production of recombinant protein therapeutics in cultivated mammalian cells. Nat Biotech. 2004;22:1393–1398. [DOI] [PubMed] [Google Scholar]
- 104. Amit M, Carpenter MK, Inokuma MS, et al. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev Biol. 2000;227:271–278. [DOI] [PubMed] [Google Scholar]
- 105. Ludwig TE, Levenstein ME, Jones JM, et al. Derivation of human embryonic stem cells in defined conditions. Nat Biotechnol. 2006;24:185–187. [DOI] [PubMed] [Google Scholar]
- 106. Yao S, Chen S, Clark J, et al. Long‐term self‐renewal and directed differentiation of human embryonic stem cells in chemically defined conditions. Proc Natl Acad Sci USA. 2006;103:6907–6912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Furue MK, Na J, Jackson JP, et al. Heparin promotes the growth of human embryonic stem cells in a defined serum‐free medium. Proc Natl Acad Sci USA. 2008;105:13409–13414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Rajala K, Lindroos B, Hussein SM, et al. A defined and xeno‐free culture method enabling the establishment of clinical‐grade human embryonic, induced pluripotent and adipose stem cells. PLoS ONE. 2010;5:e10246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Chen G, Gulbranson DR, Hou Z, et al. Chemically defined conditions for human iPSC derivation and culture. Nat Methods. 2011;8:424–429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Hasegawa K, Yasuda S, Teo J, et al. Wnt signaling orchestration with a small molecule DYRK inhibitor provides long‐term xeno‐free human pluripotent cell expansion. Stem Cells Transl Med. 2012;1:18–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Nakagawa M, Taniguchi Y, Senda S, et al. A novel efficient feeder‐free culture system for the derivation of human induced pluripotent stem cells. Sci Rep. 2014;4:3594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Hasegawa K, Yasuda S, Shahsavarani H, Yoshida N. Culture medium for pluripotent stem cells. International Patent Application, 2015, WO 2015/147047 A1.
- 113. Thibault C, Dauzier L, Wintenberger S. Cytological study of fecundation in vitro of rabbit ovum. CR Seances Soc Biol Fil. 1954;148:789–790. [PubMed] [Google Scholar]
- 114. Chang MC. Fertilization of rabbit ova in vitro. Nature. 1959;184:466–467. [DOI] [PubMed] [Google Scholar]
- 115. Whitten WK, Biggers JD. Complete development in vitro of the pre‐implantation stages of the mouse in a simple chemically defined medium. J Reprod Fertil. 1968;17:399–401. [DOI] [PubMed] [Google Scholar]
- 116. Yanagimachi R, Chang MC. Fertilization of hamster eggs in vitro. Nature. 1963;200:281–282. [DOI] [PubMed] [Google Scholar]
- 117. Steptoe PC, Edwards RG. Birth after the reimplantation of a human embryo. Lancet. 1978;2:366. [DOI] [PubMed] [Google Scholar]
- 118. Edwards RG, Steptoe PC, Purdy JM. Establishing full‐term human pregnancies using cleaving embryos grown in vitro. Br J Obstet Gynaecol. 1980;87:737–756. [DOI] [PubMed] [Google Scholar]
- 119. Noda Y. Embryo development in vitro. Assist Reprod Rev. 1992;2:9–15. [Google Scholar]
- 120. Goto Y, Noda Y, Mori T, Nakano M. Increased generation of reactive oxygen species in embryos cultured in vitro. Free Radic Biol Med. 1993;15:69–75. [DOI] [PubMed] [Google Scholar]
- 121. Bastias M, McGee‐Belser S, Bryan S, Vasquez J. In vitro deleterious effect of hypoxanthine in Ham's nutrient mixture F‐10 culture medium on human oocyte fertilization and early embryonic development. Fertil Steril. 1993;60:876–880. [DOI] [PubMed] [Google Scholar]
- 122. Loutradis D, Kiessling A, Kallianidis K, et al. A preliminary trial of human zygote culture in Ham's F‐10 without hypoxanthine. J Assist Reprod Genet. 1993;10:271–275. [DOI] [PubMed] [Google Scholar]
- 123. Quinn P, Kerin JF, Warnes GM. Improved pregnancy rate in human in vitro fertilization with the use of a medium based on the composition of human tubal fluid. Fertil Steril. 1985;44:493–498. [DOI] [PubMed] [Google Scholar]
- 124. Yao T, Asayama Y. Human preimplantation embryo culture media: past, present, and future. J Mamm Ova Res. 2016;33:17–34. [Google Scholar]
- 125. Morbeck DE, Krisher RL, Herrick JR, Baumann NA, Matern D, Moyer T. Composition of commercial media used for human embryo culture. Fertil Steril. 2014;102:759–766. [DOI] [PubMed] [Google Scholar]
- 126. Gardner DK, Lane M. Embryo culture systems In: Trounson AO, Gardner DK, eds. Handbook of In Vitro Fertilization. New York: CRC Press; 2000:205–264. [Google Scholar]
- 127. Summers MC, Biggers JD. Chemically defined media and the culture of mammalian preimplantation embryos: historical perspective and current issues. Hum Reprod Update. 2003;9:557–582. [DOI] [PubMed] [Google Scholar]
- 128. Eisenblatter T, Psathaki K, Nitz T, Galla H, Wegener J. Cell culture media: selection and standardization In: Lehr CM, ed. Cell Culture Models of Biological Barriers In‐Vitro Test Systems for Drug Absorption and Delivery. London: Taylor & Francis; 2002:20–40. [Google Scholar]
- 129. Forte TM, Bell‐Quint JJ, Cheng F. Lipoproteins of fetal and newborn calves and adult steer: a study of developmental changes. Lipids. 1981;16:240–245. [DOI] [PubMed] [Google Scholar]
- 130. Freshney RI. Defined media and supplements In: Freshney RI, ed. Culture of Animal Cells. Hoboken, NJ: John Wiley & Sons, Inc.; 2010:99–114. [Google Scholar]
- 131. Van der Valk J, Brunner D, De Smet K, et al. Optimization of chemically defined cell culture media – replacing fetal bovine serum in mammalian in vitro methods. Toxicol In Vitro. 2010;24:1053–1063. [DOI] [PubMed] [Google Scholar]
- 132. Poole CA, Reilly HC, Flint MH. The adverse effects of HEPES, TES, and BES zwitterion buffers on the ultrastructure of cultured chick embryo epiphyseal chondrocytes. In Vitro Cell Dev Biol. 1982;18:755–765. [DOI] [PubMed] [Google Scholar]
- 133. Zigler JS Jr, Lepe‐Zuniga JL, Vistica B, Gery I. Analysis of the cytotoxic effects of light‐exposed HEPES‐containing culture medium. In Vitro Cell Dev Biol. 1985;21:282–287. [DOI] [PubMed] [Google Scholar]
- 134. Grady LH, Nonneman DJ, Rottinghaus GE, Welshons WV. pH‐Dependent cytotoxicity of contaminants of phenol red for MCF‐7 breast cancer cells. Endocrinology. 1991;129:3321–3330. [DOI] [PubMed] [Google Scholar]
- 135. Walsh‐Reitz MM, Toback FG. Phenol red inhibits growth of renal epithelial cells. Am J Physiol Renal Physiol. 1992;262:F687–F691. [DOI] [PubMed] [Google Scholar]
- 136. Mandenius CF, Brundin A. Bioprocess optimization using design‐of‐experiments methodology. Biotechnol Prog. 2008;24:1191–1203. [DOI] [PubMed] [Google Scholar]
- 137. Jones B, Nachtsheim CJ. A class of three‐level designs for definitive screening in the presence of second‐order effects. J Qual Technol. 2011;43:1–15. [Google Scholar]
- 138. Menezo Y, Testart J, Perrone D. Serum is not necessary in human in vitro fertilization, early embryo culture, and transfer. Fertil Steril. 1984;42:750–755. [DOI] [PubMed] [Google Scholar]
- 139. Quinn P, Whittingham D. Effect of fatty acids on fertilization and development of mouse embryos in vitro. J Androl. 1982;3:440–444. [Google Scholar]
- 140. Yahia Khandoker M, Nihioka M, Tsujii H. Effect of BSA binding fatty acids on mouse and rat embryo development. J Mamm Ova Res. 1995;12:113–118. [Google Scholar]
- 141. Haggarty P, Wood M, Ferguson E, et al. Fatty acid metabolism in human preimplantation embryos. Hum Reprod. 2006;21:766–773. [DOI] [PubMed] [Google Scholar]
- 142. Gay M, Carrascal M, Gorga M, Pares A, Abian J. Characterization of peptides and proteins in commercial HSA solutions. Proteomics. 2010;10:172–181. [DOI] [PubMed] [Google Scholar]
- 143. Dyrlund TF, Kirkegaard K, Poulsen ET, et al. Unconditioned commercial embryo culture media contain a large variety of non‐declared proteins: a comprehensive proteomics analysis. Hum Reprod. 2014;29:2421–2430. [DOI] [PubMed] [Google Scholar]
- 144. Takatori S, Akutsu K, Kondo F, Ishii R, Nakazawa H, Makino T. Di(2‐ethylhexyl)phthalate and mono(2‐ethylhexyl)phthalate in media for in vitro fertilization. Chemosphere. 2012;86:454–459. [DOI] [PubMed] [Google Scholar]
- 145. Kane MT. Variability in different lots of commercial bovine serum albumin affects cell multiplication and hatching of rabbit blastocysts in culture. J Reprod Fertil. 1983;69:555–558. [DOI] [PubMed] [Google Scholar]
- 146. Caro CM, Trounson A. The effect of protein on preimplantation mouse embryo development in vitro. J In Vitro Fert Embryo Transf. 1984;1:183–187. [DOI] [PubMed] [Google Scholar]
- 147. Batt PA, Gardner DK, Cameron AW. Oxygen concentration and protein source affect the development of preimplantation goat embryos in vitro. Reprod Fertil Dev. 1991;3:601–607. [DOI] [PubMed] [Google Scholar]
- 148. Leveille MC, Carnegie J, Tanphaichitr N. Effects of human sera and human serum albumin on mouse embryo culture. J Assist Reprod Genet. 1992;9:45–52. [DOI] [PubMed] [Google Scholar]
- 149. McKiernan SH, Bavister BD. Different lots of bovine serum albumin inhibit or stimulate in vitro development of hamster embryos. In Vitro Cell Dev Biol. 1992;28A:154–156. [DOI] [PubMed] [Google Scholar]
- 150. Rorie RW, Miller GF, Nasti KB, McNew RW. In vitro development of bovine embryos as affected by different lots of bovine serum albumin and citrate. Theriogenology. 1994;42:397–403. [DOI] [PubMed] [Google Scholar]
- 151. Harrison K, Sherrin D, Hawthorne T, Breen T, West G, Wilson L. Embryotoxicity of micropore filters used in liquid sterilization. J In Vitro Fert Embryo Transf. 1990;7:347–350. [DOI] [PubMed] [Google Scholar]
- 152. Quinn P, Warnes GM, Kerin JF, Kirby C. Culture factors in relation to the success of human in vitro fertilization and embryo transfer. Fertil Steril. 1984;41:202–209. [DOI] [PubMed] [Google Scholar]
- 153. Tokoro M, Fukunaga N, Yamanaka K, et al. A simple method for transportation of mouse embryos using microtubes and a warm box. PLoS ONE. 2015;10:e0138854. [DOI] [PMC free article] [PubMed] [Google Scholar]