

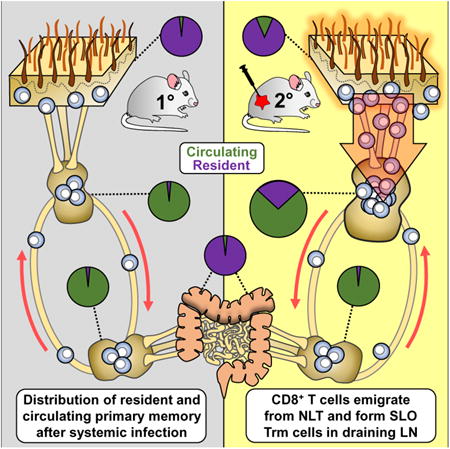
Summary
Immunosurveillance of secondary lymphoid organs (SLO) is performed by central memory T cells that recirculate through blood. Resident memory T (Trm) cells remain parked in nonlymphoid tissues and often stably express CD69. We recently identified Trm cells within SLO, but the origin and phenotype of these cells remains unclear. Using parabiosis of ‘dirty’ mice, we found that CD69 expression is insufficient to infer stable residence of SLO Trm cells. Restimulation of nonlymphoid memory CD8+ T cells within the skin or mucosa resulted in a substantial increase in bona fide Trm cells specifically within draining lymph nodes. SLO Trm cells derived from emigrants from nonlymphoid tissues and shared some transcriptional and phenotypic signatures associated with nonlymphoid Trm cells. These data indicate that nonlymphoid cells can give rise to SLO Trm cells and suggest vaccination strategies by which memory CD8+ T cell immunosurveillance can be regionalized to specific lymph nodes.
Keywords: Resident memory T cells, Secondary lymphoid organs, regionalized immunosurveillance
Graphical abstract
Introduction
Naïve T cells do not attempt to scan all 40 trillion cells in the adult human organism directly. Rather they limit immunosurveillance to secondary lymphoid organs (SLOs, including lymph nodes and splenic white pulp) that concentrate the presentation of immunogenic antigens. This surveillance pattern is achieved by SLO-selective recirculation: continuous migration between random SLOs using blood and lymphatic vessels as conduits (von Andrian and Mackay, 2000). SLO surveillance by memory T cells is also mediated by recirculation, specifically by the central memory (Tcm) cell subset that mimics naïve T cell migration patterns. In contrast, nonlymphoid tissue (NLT) immunosurveillance is dominated by nonrecirculating resident memory T cells (Trm) cells that remain parked within tissues (Mueller et al., 2013). This dichotomy supports the expectation that surveillance of SLOs, but not NLT, can be inferred by sampling blood. Moreover, residence within NLT provides a mechanism to bias memory T cell immunosurveillance to regions of the body that are most vulnerable to anamnestic infection, such as sites of repeated exposure (Schenkel and Masopust, 2014). Alternatively, SLO surveillance by memory T cells is presumed to be equilibrated among lymph nodes throughout the body.
In contrast to this model, we recently reported evidence of CD8+ Trm cells parked within SLOs of mice (Schenkel et al., 2014). Bona fide residence was demonstrated by parabiosis (Schenkel et al., 2014). Unlike their recirculating counterparts, these SLO Trm cells stably express CD69, express low levels of CD122, and persist in interleukin 15 (IL-15)-deficient mice. SLO Trm cells represent less than 5% of virus-specific memory CD8+ T cells in lymph nodes (LNs) or spleen after primary lymphocytic choriomeningitis virus (LCMV) infection. Several subsequent mouse studies support the existence of both CD4+ and CD8+ SLO Trm cells (Marriott et al., 2017; Takamura et al., 2016; Ugur et al., 2014).
CD69 is stably expressed by most Trm cells. Recent reports indicated that up to 60% of antigen-experienced CD8+ T cells isolated from the SLOs of human cadavers express CD69 (Sathaliyawala et al., 2013; Thome et al., 2014). These data raise the possibility that Trm cells make substantial contributions to immunosurveillance of human SLOs (Sathaliyawala et al., 2013; Thome et al., 2014; Woon et al., 2016). However, the interpretation is uncertain because CD69 is transiently expressed by recirculating T cells in response to antigen or inflammation, and thus may be a product of chronic infection, ephemeral responses to cytokines, or perhaps a post mortem artifact (Freeman et al., 2012; Tatovic et al., 2015; Wherry et al., 2007). Thus, unlike in vivo migration assays such as parabiosis in mice, CD69 expression alone does not provide unequivocal evidence of stable residence. The fact that many CD69+ T cells isolated from cadaverous human LNs also expressed LN entry receptors (and included naïve, stem cell memory, and Tcm subsets) is not intuitively compatible with residence, although it certainly remains possible.
Trm cells acquire a unique transcriptional program that is not shared with recirculating memory T cell subsets, and a universal Trm signature has been proposed based on analysis of CD103+ cells isolated from skin epidermis, lung, and small intestine epithelium (Mackay et al., 2013). A leading hypothesis postulates that Trm cells acquire this program in response to inductive cues encountered within nonlymphoid tissues (Casey et al., 2012; Mackay et al., 2013; Masopust et al., 2006; Skon et al., 2013). The demonstration of SLO Trm cells is seemingly incongruous with this model. This study set out to address gaps in knowledge regarding the origin and transcriptional profile of SLO Trm cells. We demonstrated that secondary antigen exposure at reproductive mucosa or skin barrier sites resulted in the accumulation of abundant virus-specific CD69+ memory CD8+ T cells specifically within the LN that drains each tissue. In vivo migration studies confirmed that these cells were indeed resident, and that they were the progeny of nonlymphoid Trm cells that gained access to the draining LN during the activation phase of the immune response. SLO Trm cells retained some, but not all, conserved features of the NLT Trm cells that have been described previously. These results reconcile the observation of SLO Trm with the model of NLT induction signals, imply epigenetic maintenance of a refined residence program, and reveal a pathway by which SLO that drain sites of repeated infection can accrue abundant local memory that is not shared with the recirculating pool or observable in blood. Imaging analyses revealed that boosting SLO Trm cells substantively increased total antigen-specific memory CD8+ T cells within all regions of specific LNs, including follicles. Lastly, using parabiosis studies in non-specific-pathogen-free (SPF) mice, we advocated caution in using CD69 expression as a sole criterion to determine stable residence, as many CD69+ CD8+ T cells (particularly those that were also CD62L+) equilibrated between mice.
Results
CD69 expression is insufficient to infer residence
CD69+ CD8+ T cells are abundant in LNs from human cadavers, but scarce in SPF mice. A small frequency of virus-specific memory CD8+ T cells express CD69 in mouse LNs after clearance of lymphocytic choriomeningitis virus (LCMV) Armstrong infection, suggesting that SLO Trm cells represent a rare population in mice (Schenkel et al., 2014). The paucity of CD69+ CD8+ T cells in murine LNs could be due to the fact that mouse LNs were freshly isolated (e.g. human data are an artifact of a cadaverous source), genetic differences between mouse and humans, or the fact that SPF laboratory mice have considerably less microbial experience than humans. To attempt to discriminate between these possibilities, we compared CD69 expression by LN CD8+ T cells among SPF inbred C57Bl/6 mice, pet store mice with diverse genetics and microbial experience, and inbred C57Bl/6 mice that acquire microbial experience via co-housing with pet store mice for >2 months, as recently described (Beura et al., 2016).
Figure 1A demonstrates that few CD8+ T cells expressed CD69 in SPF C57Bl/6 mouse freshly isolated LNs. In contrast, in pet store mice, CD69 was expressed by about 50% of the CD44hi (antigen-experienced) population and 30% of total LN CD8+ T cells, including those that expressed the LN entry receptor CD62L (Fig. 1A-B). Co-housing resulted in a similar accumulation in C57Bl/6 mice suggesting that the difference in CD69 expression among SPF mouse and human LN CD8+ T cells was largely driven by microbial experience rather than genetics or a post mortem artifact.
Figure 1. CD69 expression is insufficient to infer residence.

(A) LN CD8+ T cell phenotypes were compared between specific-pathogen free (SPF) mice, mice obtained from pet stores (Pet store) and SPF mice cohoused for at least 60 days with pet store partners (Cohoused). Representative flow plots from three independent experiments (n=3 mice per group/repeat) are shown. Plots are gated on CD8+ T lymphocytes isolated from pooled inguinal and mesenteric LNs. (B) The frequency of CD69+ cells among the indicated CD8+ T cell subsets isolated from LNs of SPF, Pet store and Cohoused mice. Data are representative of three independent experiments with 3 mice/group per experiment. (C) Experimental scheme for testing residence of LN CD8+ T cells in Cohoused mice. (D) Blood and LNs in each parabiont were analyzed by flow cytometry 15-23 days post-parabiosis for the presence of partner and host-derived cells within each indicated CD8+ T cell subset. (E) The phenotype of host and partner LN CD44lo and CD44hi CD8+ T cells. (F) The frequency of CD69+ cells in each indicated CD8+ T cell subset in LNs of parabiotic pairs. Data are representative of three independent experiments with 3 parabiotic pairs per experiment (for D, E & F). ns= not significant, ** p<0.01, **** p<0.0001, Two-way ANOVA with Sidak's multiple comparison test (for B & F). Bars indicate mean ± S.E.M.
We next asked the degree to which CD69 expression was unequivocally correlated with residence. Parabiotic surgery involves the conjoining of vasculature between two inbred mice. It is a test of residence, as recirculating cells that migrate via blood will equilibrate between each parabiont, whereas Trm cells will not. 15-23 days after parabiotic surgery of CD45.1+ and CD45.2+ co-housed C57Bl/6 mice, we observed substantial mixing between LN CD8+ T cells (Fig. 1C-D), including among CD69+ cells (Fig. 1E). Further characterization revealed that moderate disequilibrium was maintained among CD62Llo CD69+ CD8+ T cells, whereas CD62Lhi CD69+ CD8+ T cells of both CD45.1+ or CD45.2+ origin was present in similar ratios in each parabiont (Fig. 1E-F). These data suggest that some CD8 T cells may have transiently upregulated CD69 upon migration to LN in co-housed mice. Regardless, these data invite caution in interpreting residence in non-SPF organisms solely based on CD69 expression.
LN Trm cells are broadly distributed after LCMV infection
To assess LN Trm cells that do not recirculate, we turned our attention to an antigen specific model in which SLO Trm cells had previously been defined. To this end, we transferred naïve LCMV-specific CD45.1+ P14 TCR transgenic CD8+ T cells into naïve SPF C57Bl/6J mice, which were then infected with LCMV (Armstrong strain) i.p. These mice will be referred to as LCMV immune chimeras. Intraperitoneal LCMV infection has broad tropism, but is cleared within approximately one week (Beura et al., 2015). Forty five days later, lymphocytes were isolated from the indicated LNs and LCMV-specific P14 memory CD8+ T cells were assessed for expression of CD69 and CD62L (Fig. 2A). P14 memory CD8+ T cells isolated from the small intestine epithelium, spleen, and blood are shown for comparison. These data demonstrate that 1) a portion of LCMV-specific memory CD8+ T cells within all tested SLOs expressed CD69 long after detectable viral clearance (this was also true among endogenous LCMV-specific CD8+ T cells, see Fig. S1), 2) the fraction of CD69+ cells varied considerably among SLOs, and 3) CD69+ cells typically did not express CD62L in SPF mice (Fig. 2A-B). To interrogate the extent to which SLO P14 cells were stably resident, we performed parabiosis between LCMV immune chimeras containing CD90.1+ memory P14 and naïve mice that lacked P14 cells. Lymphocytes were harvested from LNs of both mice 30 days later, and the phenotype of memory CD90.1+ P14 cells in both host and naïve partner SLOs was evaluated. While CD69- cells equilibrated between hosts, CD69+CD62Llo P14 cells were absent from partner mice, indicating that this phenotype defined SLO Trm cells in this model of acute viral infection in SPF mice (Fig. 2C-D). In conclusion, after clearance of LCMV Armstrong infection, CD69+ CD8+ T cells remained in all SLOs examined, the proportions of CD69+ and CD69- LCMV-specific CD8+ T cells varied among different SLOs, and CD69 expression correlated with residence.
Figure 2. LN Trm cells are broadly distributed after LCMV infection.

(A&B) The phenotype of memory P14 CD8+ T cells isolated from the indicated location 45 days after LCMV infection. Representative data shown, 4 mice/group per experiment. (C&D) CD45.1+ P14 immune chimeric mice (host) were surgically attached to naïve CD45.2+ mice (partner). Four weeks post-parabiosis, the phenotype of P14 CD8+ T cells were measured in both host and partner parabionts. Representative data shown, two experiments with 4 parabiotic pairs per experiment (for C & D). * p<0.05, **** p<0.0001, Two-way ANOVA with Sidak's multiple comparison test (for D). Bars indicate mean ± S.E.M. See also figure S1.
Nonlymphoid tissue restimulation amplifies regionalized LN Trm cells
Figure 2 demonstrated that the proportion of Trm cells varied considerably between different SLOs, but gave no clues to their ontogeny or origin. As residence provides a mechanism for compartmentalizing immunity, we asked if local restimulation could result in selective amplification of Trm cells in regional LNs. LCMV immune chimeras were generated, which contain P14 Trm cells within the female reproductive tract (FRT) (Steinert et al., 2015). P14 were restimulated by depositing gp33 peptide into the cervical lumen as described (Schenkel et al., 2013). Ovalbumin-derived SIINFEKL peptide, which does not restimulate P14, served as a negative control. When mice were examined 30-45 days after gp33 restimulation, CD69+ Trm cells were significantly increased within the FRT draining (iliac), but not the non-draining (cervical) LNs (Fig. 3A-C). Parabiosis experiments confirmed that these cells were indeed resident (Fig. 3D-F). Importantly, intravenous transfer of recirculating memory CD8+ T cells into naïve mice, followed by t.c. antigen challenge, resulted in very few CD69+ memory cells within draining LN (see Fig. S2).
Figure 3. Nonlymphoid tissue restimulation amplifies regionalized LN Trm cells.

(A-C) LCMV immune chimeric mice were challenged with gp33 peptide transcervically (t.c.). 45 days later, the phenotype of P14 memory CD8+ T cells was compared between FRT draining (iliac) and non-draining (cervical) LNs. Data are representative of two independent experiments with 5 mice/group per experiment. (D-F) LCMV immune chimeras that were rechallenged with gp33 (or SIINFEKL control peptide) t.c. (as in A) were conjoined to naïve mice. 30 days later, FRT draining (iliac) and non-draining (cervical) LNs were interrogated for the presence and phenotype of P14 CD8+ T cells. Data are representative of two separate experiments with 4 parabiosis pairs/per experiment. (G-I) VSV-ova OT-I immune chimeric mice were challenged with SIINFEKL on the left skin flank. 30-45 days later, contralateral (non-draining) and ipsilateral (draining) LNs were interrogated for the presence and phenotype of OT-I CD8+ T cells. ns= not significant, * p<0.05, **** p<0.0001, Two-way ANOVA with Sidak's multiple comparison test (for C & F). Wilcoxon matched-pairs signed rank test (for I). Bars indicate mean ± S.E.M. See also figure S2 and S3.
We next assessed whether restimulation at an alternative site would also lead to accumulation of LN Trm specifically within regional LNs. We generated CD45.1+ OT-I (CD8+ TCR transgenic mice specific for ovalbumin-derived SIINFEKL peptide) immune chimeras through immunization with recombinant ovalbumin-expressing vesicular stomatitis virus (VSVova) infection i.v., which results in OT-I Trm cells within skin (Fig. 3G and S3). At least 30 days later, OT-I-activating SIINFEKL peptide was tattooed onto the left flank. Ipsilateral (left, draining) and contralateral (right, nondraining) inguinal LNs were evaluated for accumulation of CD69+ LN Trm cells 30-45 days after skin restimulation. Accumulation of CD69+CD62Llo P14 CD8+ T cells was highly selective for the left flank draining LN (Fig. 3H-I and data not shown). These data reveal that local restimulation at nonlymphoid sites that contain Trm cells results in a pronounced amplification of Trm cells that patrol the draining LN, but not distant LNs.
Local restimulation increases memory CD8+ T cells within all regions of the draining LN
We next tested whether local restimulation impacted the total number of antigen-specific memory CD8+ T cells specifically within draining LNs. We first generated CD45.1+ OT-I immune chimeras through i.v. immunization with VSVova, and rechallenged the left flank with SIINFEKL (as in Fig. 3G). Thirty days later, we enumerated OT-I cells within contralateral (right) and ipsilateral (left) LNs by quantitative immunofluorescence microscopy (QIM) (Steinert et al., 2015) (Fig. 4A). We found that the relative abundance of OT-I CD8+ T cells was significantly increased within the draining LN; the population more than doubled (Fig. 4B). Closer examination revealed that each region of the LN, the follicles, paracortex, sinuses, and interfollicular area, all exhibited a similar increase in the density of memory OT-I CD8+ T cells specifically within the draining LN (Fig. 4B-C). These data indicate that the augmentation of SLO Trm cells, via administration of recall antigen to local nonlymphoid tissue, can substantively increase total immunosurveillance within all compartments of regional LNs.
Figure 4. Local restimulation increases memory CD8+ T cells within all regions of draining LN.

VSV-ova OT-I immune chimeric mice were challenged with SIINFEKL peptide in the left flank (as in figure 3G). 30 days later, contralateral (right inguinal) and ipsilateral (left inguinal) LNs were examined. (A) Representative Immunofluorescence staining of frozen LN sections (scale bars, 200 μm; CD169, yellow; B220, cyan; OT-I, red). (B) Enlarged view of LNs shown in A (scale bars, 50 μm; follicle border marked with white dotted lines). (C) OT-I CD8+ T cell density in various areas of contralateral and ipsilateral inguinal LNs was analyzed by quantitative immunofluorescence microscopy. The fold increase in the abundance of OT-I CD8+ T cells in the ipsilateral LN compared to its contralateral counterpart is indicated on the top of each graph. Data are representative of two separate experiments with four mice per experiment. * p<0.05. Wilcoxon matched-pairs signed rank test.
SLO Trm cells share some phenotypic signatures with nonlymphoid Trm cells
The phenotype of Trm cells have been defined in various nonlymphoid organs, but remain much less characterized within SLOs. To address this gap, we compared the phenotype of P14 Trm cells occupying mesenteric LN and the small intestine epithelium (SI-IEL) by flow cytometry in LCMV immune chimeras generated in SPF mice. Tcm cells were also analyzed as a basis of comparison. We found that SLO Trm cells were distinct from SI Trm cells in the expression of CD103, CXCR3, and CD44 (Fig. 5A-B). However, SLO Trm cells shared signatures in common with SI Trm cells (that also distinguished them from Tcm cell), including reduced expression levels of CD122, CD27, and Ly6C. It should be noted that expression levels of Ly6C varies on Trm cells depending on location, and that this marker does not translate to humans or many mouse strains (data not shown).
Figure 5. SLO Trm cells share some phenotypic signatures with nonlymphoid Trm cells.

Sixty days after infection, P14 CD8+ T cells were isolated from LCMV immune chimeras and examined by flow cytometry. (A) Histograms comparing Tcm cells and CD69+ CD62Llo (SLO Trm) cells isolated from mesenteric LN to Trm cells isolated from the small intestine epithelium. (B) tSNE maps of the overlay of P14 Trm cells isolated from each indicated NLT (small intestine, female reproductive tract, or skin) on the Tcm, Tem, and SLO Trm cells isolated from each respective draining LN (mesenteric, iliac, or inguinal). Data are representative of two separate experiments with four mice per experiment.
T-Distributed Stochastic Neighbor Embedding (tSNE) is an algorithm for unsupervised nonlinear dimensionality reduction that compares similarities of data points in high dimensional space, and plots them in two-dimensional space. Cells that are similar cluster closer together. We applied tSNE to our poly-phenotypic flow cytometric analysis of Trm cells from three distinct nonlymphoid organs (SI, FRT, and skin), and the Tcm cells, Tem cells, and SLO Trm cells isolated from each respective NLT draining LN (either mesenteric, iliac, or inguinal LN) on a panel including 11 phenotypic markers: CD103, CD122, Ly6C, CD27, CD62L, CXCR3, CCR9, KLRG1, CX3CR1, CD69 and CD44. For each NLT and associated draining LN, SLO Trm cells clustered close to NLT Trm cells (Fig. 5C). However, they comprised a distinct population. These data further argue against overgeneralizing perceived unifying Trm signatures (Steinert et al., 2015) by demonstrating that Trm phenotype can be quite heterogeneous. For instance, just as CD69 may not predict stable residence (Fig. 1), the absence of CD103 does not predict recirculation (Fig.5A). However, it may be possible to define a truly generalizable signature that reliably predicts stable residence. To contribute to that goal, as well as providing more in depth molecular characterization of SLO Trm cells, we performed transcriptional profiling.
SLO Trm cells expressed some gene sets in common with nonlymphoid Trm cells and others that were shared with Tcm cells
Seventy days after LCMV infection, we sorted CD69-CD62Lhi P14 Tcm cells and CD69+CD62Llo SLO P14 Trm cells from the spleens (chosen based on technical considerations because spleen contained more total P14 than individual LNs) and CD69+C62LloCD103lo NLT P14 Trm cells from the FRT of LCMV immune chimeras. Gene expression patterns were analyzed between these three distinct (Tcm, SLO Trm, FRT Trm) memory T cell subsets. We compared our dataset to a previously described transcriptional signature of residence shared by CD8+ Trm cells isolated from lung, skin, and the small intestine (Mackay et al., 2013). Due to cross-platform discrepancies between probe sets, we could not validate all 37 reported Trm genes. Nevertheless, and as expected, interrogated gene expression from our FRT Trm analysis was largely in accordance with this previously reported signature (Fig. 6A). SLO Trm cells shared a restricted set of NLT Trm genes, notably those involved in trafficking and differentiation (e.g., S1pr1, S1pr5, Eomes, Itga1, and Xcl1) (Fig. 6A). For some genes, SLO Trm cells more closely aligned with Tcm cells, including low expression of immune checkpoint genes Icos and Ctla4. Independent of the previously reported NLT Trm gene signature, we identified 335 genes which were commonly regulated in SLO and FRT Trm cells (Fig. 6B and Table S1). Enrichment analysis of these shared genes upregulated in both SLO and NLT Trm revealed an activated phenotype, consistent with these cells' putative role as early responders in an anamnestic response (data not shown). Relative expression of selected common Trm genes with known relevance for T cell differentiation, function, and migration are depicted in Fig. 6C.
Figure 6. SLO Trm cells expressed some gene sets in common with nonlymphoid Trm cells and others that were shared with Tcm cells.

The indicated P14 memory CD8+ T cell subsets were sorted from spleen or FRT 70 days after LCMV infection, and microarray analysis was performed. (A) A heatmap showing the expression of 25 of the 37 genes previously reported to be differentially expressed between Tcm and Trm isolated from various NLT (Mackay et al., 2013). (B) From our dataset, a Venn diagram of all genes differentially expressed by spleen and/or FRT Trm cells as compared to Tcm cells. (C) A pairwise comparison of the fold change in expression of selected genes in spleen Trm cells and FRT Trm cells relative to Tcm cells.
SLO Trm cells can arise from nonlymphoid T cells
We demonstrated that the local administration of recall antigen resulted in an increase in Trm cells specifically within regional LNs. We thus exploited local NLT Trm cell reactivation as an experimental approach to investigate the ontogeny of SLO Trm. Immunohistochemical analysis revealed that P14 CD8+ T cells appeared in the draining afferent lymphatic vessels shortly after local peptide reactivation (Fig. 7A). To test whether cells trafficked from NLT to the draining LN after local restimulation, we crossed OT-I mice to a Kaede transgenic background (Fig. 7B). The Kaede transgene allows transient positional labeling of cells because exposure to 405nm violet light induces irreversible photoconversion of the Kaede from green to red (Tomura et al., 2008). We first generated OT-I-Kaede immune chimeras via VSVova infection, then rechallenged mice in a shaved region of skin with SIINFEKL peptide delivered by tattoo gun (similar to Fig. 3G, see methods). 72 hours later, we exposed a 2mm diameter region of skin on the left flank to 405nm violet light for 5min. 12h later, we assessed skin, ipsilateral (draining) LN, and contralateral (non-draining LN) for the presence of photoconverted OT-I (Kaede-red) cells. Per expectations, a substantive fraction (∼50-75%) of OT-IxKaede memory CD8+ T cells isolated from flank skin had photoconverted (Fig. 7B-D). We also observed photoconverted OT-I-Kaede cells in the draining LN, but only after administration of reactivating peptide (Fig. 7B-D). This result indicates that local reactivation in skin induces migration of antigen-specific CD8+ T cells from NLT to the draining LN. We obtained similar results after photoconversion and antigen re-challenge in the FRT (see Fig. S4).
Figure 7. SLO Trm cells can arise from nonlymphoid T cells.

(A) LCMV immune chimeras were challenged with gp33 peptide t.c. Two days later, uterus was examined for the localization of CD45.1+ P14 and Lyve-1+ lymphatic vessels by immunofluorescence microscopy (P14 = cyan, Lyve-1 = red, CD8β = yellow). Scale bars= 200μm (upper), 40 μm (lower). Number of P14 cells within Lyve-1+ lymphatic vessels in the FRT between gp33 and mock challenged (Ctrl) animals are compared in the right. (B-D) VSVova immune chimeric mice were made with photoconvertible OT-I-Kaede CD8+ T cells. 40 days later, mice were challenged with SIINFEKL peptide on the left flank, then exposed to violet light. 12h later, the indicated tissues were examined for the presence of photoconverted OT-I (Kaede-red+). Data are representative of three separate experiments with 3 mice per group per experiment. (E&F) CD90.1+ P14 immune chimeric mice were treated with CD90.1 depleting antibody and then challenged with gp33 t.c. 30 days later, iliac LN was examined for the presence of P14 (in F, top row gated on CD8 T cells, bottom row gated on CD90.1+ CD8+ T cells). Data are representative of two independent experiments with 5 mice per group/experiment. (G-H) Flank skins from CD90.1+ VSVova OT-I immune chimeras were engrafted onto the upper flank of CD45.1+ VSVova OT-I immune chimeras. 30 days later, graft site and surrounding skin was tattooed with SIINFEKL peptide. 20 days later, ipsilateral and contralateral LNs were examined for the presence of skin-graft derived OT-I CD8+ T cells. Data are representative of two experiments with 4 mice/experiment. * p<0.05. Two-way ANOVA with Sidak's multiple comparison test (for C). Bars indicate mean ± S.E.M. See also figure S4 and S5.
We showed previously that i.v. injection of titrated anti-CD90.1 antibody depletes CD90.1+ P14 CD8+ T cells from blood, spleen, and LNs, while preserving most P14 Trm cells present within the FRT (Schenkel et al., 2013). We exploited this strategy to test whether NLT Trm cells could give rise to SLO Trm cells. First, we generated CD90.1+ P14 LCMV immune chimeras. 2 months later, we depleted P14 cells from blood and LNs via injection of 0.75 to 1.5 mg of anti-CD90.1 antibody (Fig. 7E). Mice were then rechallenged transcervically with gp33 peptide, and rested for at least 30 days (46 days shown in Fig. 7F). We found that local peptide challenge induced the accumulation of CD62Llo CD69+ P14 cells within the FRT-draining (iliac) LN. This is consistent with additional parabiosis studies indicating that nonlymphoid memory CD8+ T cells, very likely including NLT Trm cells (>99% of P14 CD8+ T cells within the FRT are Trm (Steinert et al., 2015)), give rise to a substantive fraction of SLO Trm cells (Fig. S5). For the latter, we surgically joined CD45.1 and CD90.1 P14 immune chimeric mice, confirmed equilibration in blood, then challenged both parabionts with gp33 peptide t.c. (Fig. S5). 30 days later we enumerated the CD69+ P14 cells of both donor and host origin in the draining node of each parabiont, which revealed a significant bias for SLO Trm cells to be of host origin, indicating derivation preferentially from pre-existing Trm cells. However, there was a minor contribution by donor cells, perhaps indicating that cells recruited transiently to NLT after local antigen challenge also contribute to the SLO Trm pool. So, these data certainly do not exclude the possibility that recirculating memory CD8+ T cells, possibly recruited transiently to NLT after local antigen challenge, could also contribute to the SLO Trm pool.
To confirm that NLT could be the origin of SLO Trm cells, we returned to the OT-I VSVova immune chimera model that exploited skin SIINFEKL tattooing. In this case, we grafted 1cm2 of flank skin from a CD90.1+ OT-I immune chimera onto the upper left flank of a CD45.1+ OT-I immune chimera (Fig. 7G). Thus, the only source of CD90.1+ OT-I memory CD8+ T cells in recipient host mice is transplanted skin. 30 days after grafting, the grafted region was tattooed with SIINFEKL peptide. 15-20 days later, the draining (left) and contralateral (right) axillary LNs were examined for the presence of donor CD90.1+ OT-I cells (Fig. 7H). Skin-derived OT-I cells were observed almost exclusively in draining LN, where many expressed CD69. These data demonstrate that nonlymphoid tissues provide a source for SLO Trm cells.
Discussion
While migration assays directly test residence, they are often not feasible. The two major markers that often substitute for perceived proof of residence are CD69 and CD103. Of note, CD69 is expressed by a surprisingly abundant fraction of CD4 and CD8 T cells isolated from human SLOs, including by T cells expressing LN entry molecules and by those that appear naïve (Sathaliyawala et al., 2013; Thome et al., 2016, 2014). In contrast, CD69+ T cells are rare within SLOs of the SPF mice, or mice experiencing a primary LCMV Armstrong infection (Schenkel et al., 2014). Co-housed ‘dirty’ C57Bl/6 mice contain a broader panoply of T cell subsets, including those that are grossly underrepresented in SPF mice compared to adult humans (Beura et al., 2016). Thus, dirty mice provide additional opportunity to interrogate T cell migration properties using approaches that are not possible in human studies. Here we showed that a substantive fraction of CD69+ cells equilibrated between congenic parabionts. This observation emphasizes that CD69 is insufficient to infer residence and highlights the need to identify more stringent and reliable Trm markers.
Including bona fide SLO Trm cells along with NLT Trm cells when interrogating gene and protein expression patterns could help accelerate the identification of universal signatures that easily identify Trm cells and distinguish them from recirculating memory T cells, lymphocytes undergoing bystander activation, or cells experiencing chronic antigen stimulation. In our study, we found that SLO and FRT Trm cells shared low mRNA expression of Klf2, S1pr1, and S1pr5, which are not easily stained for by flow cytometry and also may change transiently on non-resident T cells after activation (Cyster and Schwab, 2012; Skon et al., 2013). While these transcriptional signatures also extended to previously published NLT Trm datasets (Hombrink et al., 2017; Mackay et al., 2013; Pan et al., 2017; Wakim et al., 2012), our data support a narrowing of the differential gene expression patterns considered to encompass all Trm cells (for example, one would omit markers CD103, CTLA4, and ICOS, as they were not upregulated on SLO Trm cells relative to Tcm cells). In fact, SLO Trm cells expressed many genes in common with Tcm cells that contrasted with NLT Trm cells. Indeed, a fraction of secondary memory SLO Trm cells expressed CD62L, perhaps representing the progeny of NLT Trm cells that retained the developmental plasticity for induction of CD62L expression upon a change in location (Masopust et al., 2006).
It had been proposed that Trm differentiation is regulated by local tissue-specific inductive cues (Casey et al., 2012; Mackay et al., 2013; Masopust et al., 2006). It was thus surprising to observe that SLO Trm cells, which shared a location with Tcm cells, expressed signatures in common with their NLT Trm counterparts (Schenkel et al., 2014). Here we exploited a model of local NLT Trm reactivation as a tool to investigate the ontogeny of SLO Trm cells. Our findings may reconcile the previous ontogeny model with the more recent characterization of SLO Trm cells by providing four lines of evidence supporting that SLO Trm cells are derived from T cells previously positioned within NLT. An implication of this revised model is that SLO Trm cells are capable of retaining some Trm signatures after removal from the tissue. During a primary response, it is possible that SLO Trm cells are derived from cells positioned within NLT, but egressed via the lymphatics despite being imprinted with a Trm differentiation program, and remained trapped within the draining LN. This model suggests that SLO Trm cells are not cells that never left the site of naïve T cell priming, but rather are cells that trafficked back to LN independently of blood or high endothelial venules. Similarly, cells transiently recruited to NLT during a local rechallenge, may also contribute to the SLO Trm population. Interestingly, boosting NLT Trm cells did not augment Trm cells in spleen. Primary splenic Trm cells might be derived from vascular contiguous sites of infection or NLT Trm precursors that gained access to blood and became trapped within spleen. While experimentally arduous to address, these hypotheses remain pursuits for future study.
It will be interesting to examine what aspects of Trm differentiation are epigenetically maintained, what features require tonic environmental signals, and what properties may be reprogrammable. Moreover, the change in location from NLT to SLO may contribute to alterations in Trm homeostatic maintenance, longevity, and function. For example, it was recently shown that Trm cells within the nutrient poor environment of the skin relied on free fatty acids for their metabolic needs (Pan et al., 2017). Examining how redistribution to the draining LN might rewire Trm metabolism could provide further insight into Trm biology.
Trm provide a mechanism for the organism to bias immunosurveillance to specific regions, particularly those that experienced previous infections. This concept may be extended to SLOs. Importantly, our study implies that local restimulation within specific NLT may selectively increase the magnitude of SLO Trm populations within all regions of draining LNs. This includes lymphoid follicles, which serve as reservoirs for HIV, and increasing CD8+ T cell immunosurveillance within these regions has been identified as a relevant goal for HIV vaccine strategies (Li et al., 2016; Petrovas et al., 2017). Important questions that remain are to characterize the potentially multi-faceted role of SLO Trm cells in protective immunity. It is possible that SLO Trm cells serve to protect SLOs from secondary infection (Hickman et al., 2011; Junt et al., 2007) Or, they may help coordinate recall responses among other leukocyte subsets within SLOs (Kastenmüller et al., 2013; Sung et al., 2012). Alternatively, they may provide a source for an expandable Trm population that could preferentially repopulate upstream NLT after reinfection. These possibilities are not mutually exclusive. Nevertheless, it should be emphasized that SLO Trm cells may comprise a substantive fraction of immune memories, that distribution of specific T cell populations may be highly varied between LNs, and that this population will not be observed in blood. Lastly, it highlights that the sole mechanism of LN immunosurveillance is not recirculation, thus providing an important update on classic studies of lymphocyte migration (Gowans and Knight, 1964).
Contact for Reagent and Resource Sharing
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, David Masopust (masopust@umn.edu).
Experimental Model and Subject Details
C57BL/6J (B6) and B6.SJL-PtprcaPepcb/BoyJ (CD45.1) female mice were purchased from The Jackson Laboratory and National Cancer Institute and were maintained in specific pathogen free (SPF) conditions at the University of Minnesota. Pet store mice were purchased from various pet stores in the greater Minneapolis-St. Paul metropolitan area. Information about the age of the pet store mice was not available from the vendor. Co-housing of SPF mice with sex-matched pet store partner was performed as described (Beura et al., 2016) within the University of Minnesota BSL-3 facility. CD90.1+ P14, CD45.1+ P14 and CD45.1+ OT-I, CD90.1+ OT-I mice were fully backcrossed to C57BL/6J mice and maintained in our animal colony. Kaede B6 (Tomura et al., 2008) mice were crossed to OT-I mice to generate Kaede OT-I reporter mice. All SPF and cohoused mice included in the study were female and were used at 10–20 weeks of age. All mice were housed under standard conditions in the animal facilities and used in accordance with the Institutional Animal Care and Use Committees guidelines at the University of Minnesota.
Method Details
Adoptive transfers and infections
P14 immune chimeras were generated by transferring 5×104 P14 CD8+ T cells into naive C57BL/6J mice followed by infection with 2×105 plaque-forming units (PFU) LCMV Armstrong via intraperitoneal (i.p.) injection one day later. OT-I immune chimeras were generated by transferring 5×104 naive OT-I CD8+ T cells into C57BL/6 mice. Mice were infected with 1×106 PFU Vesicular Stomatitis Virus (VSV) expressing chicken ovalbumin (VSVova) via intravenous (i.v.) route.
Intravascular staining, lymphocyte isolation and phenotyping
An intravascular staining method was used to discriminate between cells present in the vasculature from cells in the tissue parenchyma as described previously. Animals were injected i.v. with biotin/fluorochrome-conjugated anti-CD8α through the tail vein. Three minutes' post-injection, animals were sacrificed, and tissues were harvested as described (Anderson et al., 2014). Isolation of cells from skin was done as described (Collins et al., 2016). A 1-2 cm2 area of skin was harvested after shaving. The tissue was chopped into small pieces and incubated with RPMI+5% FBS containing collagenase type-III (3mg/ml) and Dnase I (5mg/ml) at 37°C with constant shaking for 1h. After t he 1h incubation, skin pieces were further dissociated using a gentlemacs dissociator (Miltenyi Biotec) and filtered twice through a 70 μm mesh before staining. Where indicated, epidermis was separated from dermis as previously described (Beura et al., 2018). Briefly, epidermal sheets were prepared from flank skin by affixing of the epidermis to slides with an optically clear double-sided adhesive (Thorlabs). Slides were incubated in 2.5mg/ml Dispase II (Roche) in PBS for 1 h at 37 °C after which dermis was physically removed. Lymphocyte isolation from other tissues and SLOs was performed as described (Thompson et al., 2016). Isolated lymphocytes were surface-stained with indicated antibodies. See Key Resources Table for the complete list of antibodies used for flow cytometry staining. The stained samples were acquired using LSRII or LSR Fortessa flow cytometers (BD) and analyzed with FlowJo software (Treestar).
Key Resources Table.
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Alexa fluor 488 anti-CD3ε antibody (145-2C11) | eBioscience- ThermoFisher | Catalog# 53-0031-82 |
| Brilliant violet 711 anti-CD45 antibody (30F-11) | BioLegend | Catalog#103147 |
| Brilliant violet 650 anti-CD45 antibody (30F-11) | BioLegend | Catalog#103151 |
| Brilliant ultraviolet 395 anti-CD90.1 antibody (OX-7) | BD Biosciences | Catalog#740261 |
| Alexa fluor 647 anti-CD90.1 antibody (OX-7) | BioLegend | Catalog#202508 |
| Purified anti-CD90.1 antibody (His51) | eBioscience- ThermoFisher | Catalog#14-0900-81 |
| PerCP/Cy5.5 anti-CD45.1 antibody (A20) | TONBO Biosciences | Catalog#65-0453-U100 |
| FITC anti-CD45.1 antibody (A20) | TONBO Biosciences | Catalog#35-0453-U500 |
| PE/Cy7 anti-CD45.2 antibody (104) | BioLegend | Catalog#109830 |
| Purified anti-CD8α antibody (53-6.7) | eBioscience- ThermoFisher | Catalog#14-0081-85 |
| Biotinylated anti-CD8α antibody (53-6.7) | eBioscience- ThermoFisher | Catalog#13-0081-82 |
| Brilliant violet 785 anti-CD8α antibody (53-6.7) | BioLegend | Catalog#100750 |
| Alexa fluor 700 anti-CD8β antibody (YTS156.7.7) | BioLegend | Catalog#126618 |
| PE anti-CD8β antibody (YTS156.7.7) | BioLegend | Catalog#126608 |
| Alexa fluor 700 anti-CD44 antibody (IM7) | BioLegend | Catalog#103026 |
| Brilliant violet 785 anti-CD44 antibody (IM7) | BioLegend | Catalog#103059 |
| Brilliant violet 510 anti-CD44 antibody (IM7) | BioLegend | Catalog#103044 |
| Brilliant violet 605 anti-CD62L antibody (MEL-14) | BioLegend | Catalog#104438 |
| Brilliant ultraviolet 737 anti-CD62L antibody (MEL-14) | BD Biosciences | Catalog#565213 |
| Brilliant violet 421 anti-CD69 antibody (H1.2F3) | BioLegend | Catalog#104528 |
| PE-CF594 anti-CD69 antibody (H1.2F3) | BD Biosciences | Catalog#562455 |
| Brilliant violet 510 anti-CD103 antibody (M290) | BD Biosciences | Catalog#563087 |
| PE anti-CD103 antibody (2E7) | BioLegend | Catalog#121406 |
| Brilliant violet 421 anti-CD122 antibody (TM-β1) | BioLegend | Catalog#339010 |
| Brilliant violet 711 anti-Ly6C antibody (HK1.4) | BioLegend | Catalog#128037 |
| APC anti-Ly6C antibody (HK1.4) | eBioscience- ThermoFisher | Catalog#17-5932-82 |
| Brilliant violet 605 anti-Ly6C antibody (HK1.4) | BioLegend | Catalog#128036 |
| FITC anti-CD27 antibody (LG.3A10) | BioLegend | Catalog#124208 |
| Brilliant ultraviolet 395 anti-CD27 antibody (LG.3A10) | BD Biosciences | Catalog#740247 |
| Brilliant violet 650 anti-CXCR3 antibody (CXCR3-173) | BioLegend | Catalog#126531 |
| APC anti-CXCR3 antibody (CXCR3-173) | eBioscience- ThermoFisher | Catalog#17-1831-82 |
| PE anti-CCR9 antibody (CW-1.2) | BioLegend | Catalog#128710 |
| PE/Cy7 anti-KLRG1antibody (2F1) | BioLegend | Catalog#138416 |
| Brilliant violet 711 anti-KLRG1antibody (2F1) | BioLegend | Catalog#138427 |
| Brilliant violet 421 anti-CX3CR1 antibody (SA011F11) | BioLegend | Catalog#149023 |
| PE anti-CD169 antibody (3D6.112) | BioLegend | Catalog#142404 |
| Alexa fluor 647 anti-B220 antibody (RA3-6B2) | BioLegend | Catalog#103226 |
| PE anti-B220 antibody (RA3-6B2) | eBioscience- ThermoFisher | Catalog#12-0452-82 |
| Rat anti-mouse LYVE-1 antibody (223322) | R&D Systems | Catalog#MAB-2125(500) |
| Donkey anti-rat Alexa fluor 647 secondary antibody | Jackson Immunoresearch | Catalog#712-605-150 |
| Bacterial and Virus Strains | ||
| Lymphocytic choriomeningitis virus (LCMV)- Armstrong strain | Dr. R. Ahmed, Emory University | N/A |
| Vesicular stomatitis virus (VSV)- Indiana strain expressing ovalbumin (VSVova) | Dr. L. Lefrancois, Univ. of Connecticut | N/A |
| Chemicals, Peptides, and Recombinant Proteins | ||
| H-2Kb -restricted peptide SIINFEKL | New England Peptide (Custom peptide synthesis) | N/A |
| H-2Db -restricted peptide GP33-41 | New England Peptide (Custom peptide synthesis) | N/A |
| Collagenase I | Worthington Biochemicals | Catalog#LS004197 |
| Collagenase III | Worthington Biochemicals | Catalog#LS004183 |
| Collagenase IV | Sigma Aldrich | Catalog#C5138-5G |
| Dispase II | Roche biosciences | Catalog#04942078001 |
| Dnase I | Sigma Aldrich | Catalog#D4513 |
| 4′,6-Diamidino-2-Phenylindole dihydrochloride (DAPI) | Sigma Aldrich | Catalog#D9542 |
| Sytox green | ThermoFisher Scientific | Catalog#S7020 |
| Live dead Ghost Dye™ Red 780 | TONBO Biosciences | Catalog#13-0865-T500 |
| H-2Db /GP33-41 KAVYNFATM biotinylated monomer | Prepared in house following NIAID tetramer core facility protocol | N/A |
| H-2Kb /OVA257-264 SIINFEKL biotinylated monomer | Prepared in house following NIAID tetramer core facility protocol | N/A |
| Streptavidin-APC (monomer tetramerization) | Life Technologies | Catalog#S868 |
| Streptavidin-PE (monomer tetramerization) | Life Technologies | Catalog#S866 |
| Critical Commercial Assays | ||
| EasySep™ Mouse CD8+ T Cell Isolation Kit | STEMCELL Technologies | Catalog#19853 |
| RNeasy Plus Micro Kit | Qiagen | Catalog#74034 |
| GeneChip™ 3′ IVT Pico Kit | Affymetrix ThermoFisher Scientific | Catalog#902789 |
| QIAshredder columns | Qiagen | Catalog#79654 |
| Deposited Data | ||
| Transcriptional comparison between Tcm, FRT Trm and SLO Trm | This paper | GEO accession # GSE109060 |
| Experimental Models: Cell Lines | ||
| Baby hamster kidney-21 cells | ATCC | Catalog#BHK-21 [C-13] (ATCC CCL-10™) |
| Vero cells | ATCC | Catalog#Vero (ATCC CCL-81™) |
| Experimental Models: Organisms/Strains | ||
| C57BL/6J | The Jackson Laboratory | Catalog# JAX:000664, RRID: IMSR_JAX:000664 |
| C57BL/6NCrl | Charles River Laboratories | Catalog# CRL:27,RRID:IMSR_CRL:27 |
| P14 | Dr. R. Ahmed, Emory University | Catalog# MUGEN:M189001, RRID:IMSR_MUGEN:M189001 |
| OT-I | Dr. K. Hogquist, Univ. of Minnesota | Catalog# JAX:003831, RRID:IMSR_JAX:003831 |
| Kaede B6 | Dr. M. Tomura, RIKEN & Dr. K. Hogquist, Univ. of Minnesota | Catalog# RBRC05737, RRID:IMSR_RBRC05737 |
| B6 CD45.1 | Charles River Laboratories | Catalog# CRL:564, RRID:IMSR_CRL:564 |
| B6 Thy1.1 | The Jackson Laboratory | Catalog# JAX:000406, RRID:IMSR_JAX:000406 |
| Pet store mice | Pet stores in the greater Minneapolis-St. Paul metropolitan area | N/A |
| Software and Algorithms | ||
| affy | Bioconductor (Gautier et al., 2004) | RRID:SCR_012835 |
| GSEA | Broad Institute (Mootha et al. 2003) | RRID:SCR_003199 |
| LIMMA | Bioconductor (Ritchie et al. 2015) | RRID:SCR_010943 |
| GOseq | http://bioinf.wehi.edu.au/software/goseq/ (Young et al. 2010) | N/A |
| Prism 7 | Graphpad Inc | RRID:SCR_002798 |
| Flowjo v9 and v10 | Treestar Inc | RRID:SCR_008520 |
Tissue freezing, immunofluorescence and microscopy
Harvested murine tissues were fixed in 2% paraformaldehyde for 2hrs before being treated with 30% sucrose overnight for cryoprotection. The sucrose treated tissue was embedded in tissue freezing medium OCT and frozen in an isopentane liquid bath. Frozen blocks were cut in a Leica cryostat to prepare 7 μm-thick sections. Slides were stained and immunofluorescence microscopy was performed using a Leica DM6000 B microscope (Schenkel et al., 2013; Steinert et al., 2015). Counterstaining with either 4′,6-Diamidino-2-Phenylindole dihydrochloride (DAPI) (Sigma Aldrich) or sytox green (Thermofisher) was done to detect nuclei. Enumeration of various cell types was performed using ImageJ64 as described before (Schenkel et al., 2013; Steinert et al., 2015) except the QIM procedures are semi-automated. Images underwent gaussian filtering, thresholding and local maxima detection to identify individual cells. Antibodies and chemical used for staining are mentioned in the Key Resources Table.
RNA isolation and microarray hybridization
CD8+ CD90.1+ P14 Tcm (CD62Lhi CD69-) and SLO Trm (CD62Llo CD69+) populations were sorted from spleen of LCMV immune chimeras using a BD FACSAria II. For isolation of Trm cells from Female reproductive tract (FRT), tissues were cut into small pieces in RPMI 1640 containing 5% FBS, 2 mM MgCl2, 2 mM CaCl2, and 0.5 mg/ml type IV collagenase (Sigma-Aldrich) and incubated for 1 h at 37°C with stirring. After enzymatic digestion, the remaining tissue pieces were mechanically disrupted using a gentleMACs dissociator (Miltenyi Biotec, San Diego, CA). The resulting single-cell suspensions was used as a source for sorting CD69+C62LloCD103lo Trm cells from FRT. Sorted cells were homogenized using QIAshredder columns (Qiagen) and RNA was extracted using an RNeasy kit (Qiagen) per the manufacturer's instructions. RNA quality was assessed using the Agilent Model 2100 Bioanalyzer. Following quality control, total RNA samples were processed using the GeneChip® 3′ IVT Pico Kit (Affymetrix). Samples were hybridized and loaded onto Affymetrix Genechip Mouse Genome 430 2.0 arrays.
Microarray analysis
All the raw data files obtained by the Affymetrix scanner passed the data quality control steps. The probe data were then preprocessed and normalized with RMA (Robust Multiarray Average) from the “affy” R/Bioconductor package (Gautier et al., 2004). Processed probe expression was then collapsed to gene expression using the Collapse DataSet function in Gene Set Enrichment Analysis (GSEA) with the following settings: collapsing mode for probe sets ≥ 1 gene = Max_probe and Chip platforms = GENE_SYMBOL.chip (Mootha et al., 2003). Prior to performing differential expression analysis, genes with average expression across all samples less than the background expression level (expression < 3.5) were filtered out. Differential gene expression analysis was performed using the LIMMA package in R (Ritchie et al., 2015), and FDR-adjusted p-values were used to select statistically significant genes (FDR cut-off = 0.05). Gene Ontology term enrichment analysis was performed for differentially expressed gene sets using the “GOseq” package in R (Young et al., 2010).
In vivo antibody treatment, local Trm reactivation, and photoconversion
Circulating Thy1.1+ P14 CD8+ T cells were depleted by injecting 0.75 to 1.5 μg of titrated Thy1.1 (HIS51, eBioscience) antibody i.p. as previously described (Schenkel et al., 2013). For local Trm reactivation experiments, 50μg of the indicated peptides was delivered t.c. as described (Schenkel et al., 2013) in a volume of 30μl by modified gel loading pipet. To reactivate OT-I CD8+ Trm cells positioned in skin, a 2cm2 area of the flank skin was shaved and 0.5 μg of SIINFEKL peptide was applied using a tattoo gun. PBS was used in control animals. Peptide tattooing was performed on only one side of mouse (ipsilateral) and the contralateral side is treated as a control. For photoconversion of skin Kaede OT-I Trm cells, a reactivated area of skin (5 mm2) was exposed to violet light (405nm ± 5nm) using a Fiber Coupled Violet LED light source (Optogenetics-LED-Violet, Prizmatix) for 5 minutes. The tattooed and photoconversion sites were identical. The surrounding skin was covered with aluminum foil. For photoconversion of cells in the mouse uterus, the violet LED light source was modified to be connected to a 43mm long cannula (with a core diameter 500μm, NA:0.63) using zirconia mating sleeves. The cannula was gently inserted into the vagina of a sedated animal and guided through the cervical canal and stably positioned inside the uterine horn for the duration of photo exposure (5 minutes).
Parabiosis and skin transplant surgeries
Parabiosis surgery was done as described (Steinert et al., 2015). Parabiosed mice were then allowed to rest for 14–30 days before experiments. Equilibration was confirmed in the peripheral blood prior to Trm cell reactivation. Skin transplant was performed as described earlier (Silva and Sundberg, 2013). Briefly, depilated flank skin was harvested from donor CD90.1/OT-I immune chimeric mice. The recipient graft bed was prepared by excising a ∼1 cm2 piece of skin from the upper left flank. The donor skin was sutured on to the graft site using silk sutures (Sofsilk, Covidien) and a band aid was used to keep the graft in place. The band aid and sutures were removed 7-10 days' post-surgery and the graft was allowed to heal for at least 30 days before peptide challenge.
Quantification and Statistical Analysis
If the samples followed normal distribution, then parametric tests (unpaired two-tailed Student's t-test for two groups and one-way ANOVA with Tukey's multiple comparison test for more than two groups) were used. Two-way ANOVA with Sidak's multiple comparison test was used if the effect of two independent variables were being considered among more than two sample groups. If the samples deviated from a Gaussian distribution, non-parametric tests (Mann–Whitney U test for two groups, Kruskal–Wallis with Dunn's multiple comparison test for more than two groups) were used unless otherwise stated in the figure legend. For paired analyses not conforming to Gaussian distribution, Wilcoxon matched-pair signed rank test was used. D'Agostino and Pearson omnibus normality test was used to determine whether samples adhered to Gaussian distribution or not. Variances between groups were compared using an F test and found to be equal. All statistical analysis was done in GraphPad Prism (GraphPad Software Inc.). P < 0.05 was considered significant.
Data and Software Availability
Microarray data from the three different cell populations (Tcm, FRT Trm and SLO Trm cells) are available at the NCBI GEO under accession number GSE109060.
Supplementary Material
Table S1. Related to figure 6: Commonly regulated genes in FRT Trm cells and SLO Trm cells. FRT Trm, SLO Trm, and Tcm P14 memory CD8+ T cell subsets were sorted from spleen or FRT 70 days after LCMV infection, and microarray analysis was performed. 335 genes that were commonly upregulated or downregulated in FRT Trm and SLO Trm cells (as compared to Tcm) are shown. logFC: log2-expression fold change; AveExpr: average log2-expression for all experimental samples; t: moderated t-statistic; adj.P.Val: Benjamini-Hochberg corrected P-value; B: B-statistic.
Acknowledgments
We thank the members of the Masopust laboratory for helpful discussions. The authors acknowledge the assistance of the University of Minnesota Flow Cytometry core facility, the Biosafety level 3 program, Thomas Pengo (University Imaging Center) and funding by National Institutes of Health grants R01AI111671, R01AI084913 and the Howard Hughes Medical Institute Faculty Scholars program (D.M.).
Abbreviations
- Trm
Resident memory T cells
- Tcm
Central memory T cells
- Tem
Effector memory T cells
- FRT
Female reproductive tract
- QIM
Quantitative Immunofluorescence microscopy
- NLT
Non lymphoid tissue
- SLO
Secondary lymphoid organ
- SI IEL
small intestine intraepithelial lymphocyte
Footnotes
Author contribution: L.K.B., S.W., E.A.T., P.C.R., M.J.P., and J.M.S. performed the experiments; J.S.M., V.V., and B.T.F. contributed critical reagents and experimental help; M.G.M., S.W., and S.S. performed transcriptional analysis. L.K.B., S.W., and D.M. wrote the manuscript; and D.M. was responsible for research supervision, coordination and strategy.
Declaration of Interests: The authors declare no competing interests.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Anderson KG, Mayer-Barber K, Sung H, Beura L, James BR, Taylor JJ, Qunaj L, Griffith TS, Vezys V, Barber DL, Masopust D. Intravascular staining for discrimination of vascular and tissue leukocytes. Nat Protoc. 2014;9:209–22. doi: 10.1038/nprot.2014.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beura LK, Anderson KG, Schenkel JM, Locquiao JJ, Fraser KA, Vezys V, Pepper M, Masopust D. Lymphocytic choriomeningitis virus persistence promotes effector-like memory differentiation and enhances mucosal T cell distribution. J Leukoc Biol. 2015;97:217–25. doi: 10.1189/jlb.1HI0314-154R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beura LK, Hamilton SE, Bi K, Schenkel JM, Odumade OA, Casey KA, Thompson EA, Fraser KA, Rosato PC, Filali-Mouhim A, Sekaly RP, Jenkins MK, Vezys V, Haining WN, Jameson SC, Masopust D. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature. 2016;532:512–6. doi: 10.1038/nature17655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beura LK, Mitchell JS, Thompson EA, Schenkel JM, Mohammed J, Wijeyesinghe S, Fonseca R, Burbach BJ, Hickman HD, Vezys V, Fife BT, Masopust D. Intravital mucosal imaging of CD8+ resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory. Nat Immunol. 2018;19:173–182. doi: 10.1038/s41590-017-0029-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey KA, Fraser KA, Schenkel JM, Moran A, Abt MC, Beura LK, Lucas PJ, Artis D, Wherry EJ, Hogquist K, Vezys V, Masopust D. Antigen-independent differentiation and maintenance of effector-like resident memory T cells in tissues. J Immunol. 2012;188:4866–75. doi: 10.4049/jimmunol.1200402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins N, Jiang X, Zaid A, Macleod BL, Li J, Park CO, Haque A, Bedoui S, Heath WR, Mueller SN, Kupper TS, Gebhardt T, Carbone FR. Skin CD4(+) memory T cells exhibit combined cluster-mediated retention and equilibration with the circulation. Nat Commun. 2016;7:11514. doi: 10.1038/ncomms11514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cyster JG, Schwab SR. Sphingosine-1-Phosphate and Lymphocyte Egress from Lymphoid Organs. Annu Rev Immunol. 2012;30:69–94. doi: 10.1146/annurev-immunol-020711-075011. [DOI] [PubMed] [Google Scholar]
- Freeman BE, Hammarlund E, Raué HP, Slifka MK. Regulation of innate CD8+ T-cell activation mediated by cytokines. Proc Natl Acad Sci U S A. 2012;109:9971–6. doi: 10.1073/pnas.1203543109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gautier L, Cope L, Bolstad BM, Irizarry RA. Affy - Analysis of Affymetrix GeneChip data at the probe level. Bioinformatics. 2004;20:307–315. doi: 10.1093/bioinformatics/btg405. [DOI] [PubMed] [Google Scholar]
- Gowans JL, Knight EJ. The route of re-circulation of lymphocytes in rats. Proc R Soc London. 1964;159:257–282. doi: 10.1098/rspb.1964.0001. [DOI] [PubMed] [Google Scholar]
- Hickman HD, Li L, Reynoso GV, Rubin EJ, Skon CN, Mays JW, Gibbs J, Schwartz O, Bennink JR, Yewdell JW. Chemokines control naive CD8+ T cell selection of optimal lymph node antigen presenting cells. J Exp Med. 2011;208:2511–24. doi: 10.1084/jem.20102545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hombrink P, Helbig C, Backer RA, Piet B, Oja AE, Stark R, Brasser G, Jongejan A, Jonkers RE, Nota B, Basak O, Clevers HC, Moerland PD, Amsen D, van Lier RAW. Programs for the persistence, vigilance and control of human CD8+ lung-resident memory T cells. Nat Immunol. 2016;17:1467–1478. doi: 10.1038/ni.3589. [DOI] [PubMed] [Google Scholar]
- Junt T, Moseman EA, Iannacone M, Massberg S, Lang PA, Boes M, Fink K, Henrickson SE, Shayakhmetov DM, Di Paolo NC, van Rooijen N, Mempel TR, Whelan SP, von Andrian UH. Subcapsular sinus macrophages in lymph nodes clear lymph-borne viruses and present them to antiviral B cells. Nature. 2007;450:110–114. doi: 10.1038/nature06287. [DOI] [PubMed] [Google Scholar]
- Kastenmüller W, Brandes M, Wang Z, Herz J, Egen JG, Germain RN. Peripheral Prepositioning and Local CXCL9 Chemokine-Mediated Guidance Orchestrate Rapid Memory CD8+ T Cell Responses in the Lymph Node. Immunity. 2013;38:502–513. doi: 10.1016/j.immuni.2012.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Folkvord JM, Rakasz EG, Abdelaal HM, Wagstaff RK, Kovacs KJ, Kim HO, Sawahata R, MaWhinney S, Masopust D, Connick E, Skinner PJ. Simian Immunodeficiency Virus-Producing Cells in Follicles Are Partially Suppressed by CD8 + Cells In Vivo. J Virol. 2016;90:11168–11180. doi: 10.1128/JVI.01332-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay LK, Rahimpour A, Ma JZ, Collins N, Stock AT, Hafon ML, Vega-Ramos J, Lauzurica P, Mueller SN, Stefanovic T, Tscharke DC, Heath WR, Inouye M, Carbone FR, Gebhardt T. The developmental pathway for CD103(+)CD8+ tissue-resident memory T cells of skin. Nat Immunol. 2013;14:1294–301. doi: 10.1038/ni.2744. [DOI] [PubMed] [Google Scholar]
- Marriott CL, Dutton EE, Tomura M, Withers DR. Retention of Ag-specific memory CD4 + T cells in the draining lymph node indicates lymphoid tissue resident memory populations. Eur J Immunol. 2017;44 doi: 10.1002/eji.201646681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masopust D, Vezys V, Wherry EJ, Barber DL, Ahmed R. Cutting edge: gut microenvironment promotes differentiation of a unique memory CD8 T cell population. J Immunol. 2006;176:2079–83. doi: 10.4049/jimmunol.176.4.2079. [DOI] [PubMed] [Google Scholar]
- Mootha VK, Lindgren CM, Eriksson K, Subramanian A, Sihag S, Lehar J, Puigserver P, Carlsson E, Ridderstråle M, Laurila E, Houstis N, Daly MJ, Patterson N, Mesirov JP, Golub TR, Tamayo P, Spiegelman B, Lander ES, Hirschhorn JN, Altshuler D, Groop LC. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat Genet. 2003;34:267–273. doi: 10.1038/ng1180. [DOI] [PubMed] [Google Scholar]
- Mueller SN, Gebhardt T, Carbone FR, Heath WR. Memory T Cell Subsets, Migration Patterns, and Tissue Residence. Annu Rev Immunol. 2013;31:137–61. doi: 10.1146/annurev-immunol-032712-095954. [DOI] [PubMed] [Google Scholar]
- Pan Y, Tian T, Park CO, Lofftus SY, Mei S, Liu X, Luo C, O'Malley JT, Gehad A, Teague JE, Divito SJ, Fuhlbrigge R, Puigserver P, Krueger JG, Hotamisligil GS, Clark RA, Kupper TS. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism. Nature. 2017;543:252–256. doi: 10.1038/nature21379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrovas C, Ferrando-Martinez S, Gerner MY, Casazza JP, Pegu A, Deleage C, Cooper A, Hataye J, Andrews S, Ambrozak D, Del Río Estrada PM, Boritz E, Paris R, Moysi E, Boswell KL, Ruiz-Mateos E, Vagios I, Leal M, Ablanedo-Terrazas Y, Rivero A, Gonzalez-Hernandez LA, McDermott AB, Moir S, Reyes-Terán G, Docobo F, Pantaleo G, Douek DC, Betts MR, Estes JD, Germain RN, Mascola JR, Koup RA. Follicular CD8 T cells accumulate in HIV infection and can kill infected cells in vitro via bispecific antibodies. Sci Transl Med. 2017;9:eaag2285. doi: 10.1126/scitranslmed.aag2285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, Smyth GK. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015;43:e47. doi: 10.1093/nar/gkv007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sathaliyawala T, Kubota M, Yudanin N, Turner D, Camp P, Thome JJC, Bickham KL, Lerner H, Goldstein M, Sykes M, Kato T, Farber DL. Distribution and Compartmentalization of Human Circulating and Tissue-Resident Memory T Cell Subsets. Immunity. 2013;38:187–197. doi: 10.1016/j.immuni.2012.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenkel JM, Fraser KA, Masopust D. Cutting edge: resident memory CD8 T cells occupy frontline niches in secondary lymphoid organs. J Immunol. 2014;192:2961–4. doi: 10.4049/jimmunol.1400003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenkel JM, Fraser KA, Vezys V, Masopust D. Sensing and alarm function of resident memory CD8(+) T cells. Nat Immunol. 2013;14:509–513. doi: 10.1038/ni.2568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenkel JM, Masopust D. Tissue-resident memory T cells. Immunity. 2014;41:886–897. doi: 10.1016/j.immuni.2014.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva KA, Sundberg JP. Surgical Methods for Full-Thickness Skin Grafts to Induce Alopecia Areata in C3H / HeJ Mice. Comp Med. 2013;63:392–397. [PMC free article] [PubMed] [Google Scholar]
- Skon CN, Lee JY, Anderson KG, Masopust D, Hogquist KA, Jameson SC. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+ T cells. Nat Immunol. 2013;14:1285–93. doi: 10.1038/ni.2745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinert EM, Schenkel JM, Fraser KA, Beura LK, Manlove LS, Igyártó BZ, Southern PJ, Masopust D. Quantifying memory CD8 T cells reveals regionalization of immunosurveillance. Cell. 2015;161:737–749. doi: 10.1016/j.cell.2015.03.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sung JH, Zhang H, Ashley Moseman E, Alvarez D, Iannacone M, Henrickson SE, De La Torre JC, Groom JR, Luster AD, Von Andrian UH. Chemokine guidance of central memory T cells is critical for antiviral recall responses in lymph nodes. Cell. 2012;150:1249–1263. doi: 10.1016/j.cell.2012.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takamura S, Yagi H, Hakata Y, Motozono C, McMaster SR, Masumoto T, Fujisawa M, Chikaishi T, Komeda J, Itoh J, Umemura M, Kyusai A, Tomura M, Nakayama T, Woodland DL, Kohlmeier JE, Miyazawa M. Specific niches for lung-resident memory CD8+ T cells at the site of tissue regeneration enable CD69-independent maintenance. J Exp Med. 2016:1–17. doi: 10.1084/jem.20160938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatovic D, Young P, Kochba E, Levin Y, Wong FS, Dayan CM. Fine-Needle Aspiration Biopsy of the Lymph Node: A Novel Tool for the Monitoring of Immune Responses after Skin Antigen Delivery. J Immunol. 2015;195:386–392. doi: 10.4049/jimmunol.1500364. [DOI] [PubMed] [Google Scholar]
- Thome JJ, Grinshpun B, Kumar BV, Kubota M, Ohmura Y, Lerner H, Sempowski GD, Shen Y, Farber DL. Longterm maintenance of human naive T cells through in situ homeostasis in lymphoid tissue sites 6506. 2016:1–12. doi: 10.1126/sciimmunol.aah6506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thome JJC, Yudanin N, Ohmura Y, Kubota M, Grinshpun B, Sathaliyawala T, Kato T, Lerner H, Shen Y, Farber DL. Spatial map of human t cell compartmentalization and maintenance over decades of life. Cell. 2014;159:814–828. doi: 10.1016/j.cell.2014.10.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson Ea, Beura LK, Nelson CE, Anderson KG, Vezys V. Shortened Intervals during Heterologous Boosting Preserve Memory CD8 T Cell Function but Compromise Longevity. J Immunol. 2016;196:3054–63. doi: 10.4049/jimmunol.1501797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomura M, Yoshida N, Tanaka J, Karasawa S, Miwa Y, Miyawaki A, Kanagawa O. Monitoring cellular movement in vivo with photoconvertible fluorescence protein “Kaede” transgenic mice. Proc Natl Acad Sci U S A. 2008;105:10871–10876. doi: 10.1073/pnas.0802278105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ugur M, Schulz O, Menon MB, Krueger A, Pabst O. Resident CD4+ T cells accumulate in lymphoid organs after prolonged antigen exposure. Nat Commun. 2014;5:4821. doi: 10.1038/ncomms5821. [DOI] [PubMed] [Google Scholar]
- von Andrian UH, Mackay CR. T-cell function and migration. Two sides of the same coin. N Engl J Med. 2000;343:1020–1034. doi: 10.1056/NEJM200010053431407. [DOI] [PubMed] [Google Scholar]
- Wakim LM, Woodward-Davis A, Liu R, Hu Y, Villadangos J, Smyth G, Bevan MJ. The molecular signature of tissue resident memory CD8 T cells isolated from the brain. J Immunol. 2012;189:3462–71. doi: 10.4049/jimmunol.1201305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wherry EJ, Ha SJ, Kaech SM, Haining WN, Sarkar S, Kalia V, Subramaniam S, Blattman JN, Barber DL, Ahmed R. Molecular Signature of CD8+ T Cell Exhaustion during Chronic Viral Infection. Immunity. 2007;27:670–684. doi: 10.1016/j.immuni.2007.09.006. [DOI] [PubMed] [Google Scholar]
- Woon HG, Braun A, Li J, Smith C, Edwards J, Sierro F, Feng CG, Khanna R, Elliot M, Bell A, Hislop AD, Tangye SG, Rickinson AB, Gebhardt T, Britton WJ, Palendira U. Compartmentalization of Total and Virus-Specific Tissue-Resident Memory CD8+ T Cells in Human Lymphoid Organs. PLoS Pathog. 2016;12:1–19. doi: 10.1371/journal.ppat.1005799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young MD, Wakefield MJ, Smyth GK, Oshlack A. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 2010;11:R14. doi: 10.1186/gb-2010-11-2-r14. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Related to figure 6: Commonly regulated genes in FRT Trm cells and SLO Trm cells. FRT Trm, SLO Trm, and Tcm P14 memory CD8+ T cell subsets were sorted from spleen or FRT 70 days after LCMV infection, and microarray analysis was performed. 335 genes that were commonly upregulated or downregulated in FRT Trm and SLO Trm cells (as compared to Tcm) are shown. logFC: log2-expression fold change; AveExpr: average log2-expression for all experimental samples; t: moderated t-statistic; adj.P.Val: Benjamini-Hochberg corrected P-value; B: B-statistic.