

SUMMARY
Rational immunogen design aims to focus antibody responses to vulnerable sites on primary antigens. Given the size of these antigens, there is, however, potential for eliciting unwanted, off-target responses. Here, we use our electron microscopy polyclonal epitope mapping approach to describe the antibody specificities elicited by immunization of non-human primates with soluble HIV envelope trimers and subsequent repeated viral challenge. An increased diversity of epitopes recognized and the approach angle by which these antibodies bind constitute a hallmark of the humoral response in most protected animals. We also show that fusion peptide-specific antibodies are likely responsible for some neutralization breadth. Moreover, cryoelectron microscopy (cryo-EM) analysis of a fully protected animal reveals a high degree of clonality within a subset of putatively neutralizing antibodies, enabling a detailed molecular description of the antibody paratope. Our results provide important insights into the immune response against a vaccine candidate that entered into clinical trials in 2019.
Graphical Abstract

In Brief
Nogal et al. use electron microscopy polyclonal epitope mapping of BG505 Env-immunized and matched SHIVBG505-challenged non-human primates to identify hallmarks of protection. Additionally, cryo-EM polyclonal analysis of a fully protected animal reveals a high degree of clonality, allowing detailed characterization of a putative neutralizing paratope.
INTRODUCTION
Despite marked progress toward HIV management with effective anti-retroviral therapy (ART) options, HIV and acquired immunodeficiency syndrome (AIDS) remain major global health challenges. Although ART and other prevention options have reduced the overall incidence of HIV/AIDS (Cohen et al., 2011), nearly a million individuals are newly infected each year. Thus, a vaccine that protects from infection remains the most robust approach for eradicating HIV across diverse populations. However, due in no small part to the high rate of mutation of HIV and the associated mastery of disguise by a dense and adaptable glycan shield, efforts toward developing an immunogen that would impart protection from global HIV subtypes have thus far failed. Although correlates and mechanisms of vaccine protection are still being confirmed in clinical trials, the passive transfer of neutralizing antibodies (nAbs) in rhesus macaques has shown to reproducibly protect against simian-HIV (SHIV) challenge (Ahmed et al., 2017; Burton and Hangartner, 2016; Lu et al., 2016). Thus, broadly neutralizing antibodies (bnAbs) and the associated epitopes that they engage on the HIV envelope protein (Env) have helped to inform HIV rational vaccine design, with the ultimate goal of bnAb elicitation and thus protection from global HIV subtypes via immunization.
HIV bnAbs are present in a minority of HIV-1-infected individuals and are present at relatively rare frequencies in the overall humoral response to infection (Cohen and Frahm, 2017). The epitopes targeted by bnAbs include the CD4 binding site (CD4bs), the variable region 1 and 2 glycan site (V1/V2-glycan), the N332 supersite, the glycoprotein 120 (gp120)-gp41 interface, the silent face (VRC-PG05) (Zhou et al., 2018), and the membrane proximal external region (MPER) (Burton and Hangartner, 2016). bnAbs generally have atypical features compared with the normal antibody repertoire, including unusually long heavy chain complementary determining region 3 (CDRH3) or short CDRL3s, auto- and poly-reactivity, and/or high levels of somatic hypermutation (SHM) (West et al., 2014; Yu and Guan, 2014). Moreover, many germline-reverted bnAbs have little or no affinity for the HIV envelope trimer (Escolano et al., 2016; Prabakaran et al., 2014). Thus, elicitation of bnAbs within the practical framework of a prime-boost vaccination approach is not trivial. To date, human clinical trials have shown no or limited impact, with only one, based on gp120, showing limited efficacy (Haynes et al., 2016; Hsu and O’Connell, 2017; Rerks-Ngarm et al., 2009). Recent animal studies using novel immunogens designed to present a more native structure and antigenic surface of the Env trimer are, however, defining a new and potentially promising path toward eliciting more productive immune responses (Jardine et al., 2013).
One such protein subunit immunogen is the BG505 SOSIP.664 gp140 trimer, a mimic of the natural Env spike protein, that is stabilized in a prefusion conformation, which is a state recognized by broadly nAbs, but not non-nAbs (Chuang et al., 2017). This immunogen recapitulates the quaternary structural features and surface glycans that Abs must learn to navigate as they mature into potent and broad responses via SHM. As this vaccine candidate enters human clinical trials (ClinicalTrials.gov: NCT03699241), an examination of elicited antibody response in an animal model that more closely approximates the complexity of the human immune system is timely. Pauthner et al. (2017) recently reported success in eliciting protective tier-2 virus neutralization responses by BG505 SOSOP.664 immunization in rhesus macaques. Immunized non-human primates (NHPs) were grouped into “high-titer” or “low-titer” animals, defined in the Pauthner et al. (2017, 2019) studies, depending on the ability of their serum antibodies to neutralize BG505 pseudo-virus. Both groups, matched with non-immunized controls, were then repetitively challenged with a chimeric SHIV expressing the autologous BG505 Env (SHIVBG505) (Pauthner et al., 2019). Because the variable levels of both neutralization titers and protection from challenge observed in these macaques may hold important clues as to the potentially productive and unproductive immune system paths that may be evoked by the native-like SOSIP trimer, we decided to perform an analysis of epitopes recognized in the animals from these groups, and to compare them with the antibody response elicited following infection of non-immunized animals.
Despite its native antigenic profile, the BG505 SOSIP.664 trimer exposes surfaces such as the base of the trimer that are not accessible on the full-length, membrane-inserted Env protein and that have been shown to be very immunogenic in the rabbits (Bianchi et al., 2018). Whereas humoral responses against the base of the trimer are irrelevant, the responses to the other regions of the trimer have the ability to elicit protective tier-2 nAbs. However, these responses typically comprise variable epitopes that are strain specific with little to no potential for developing breadth. Further, these unwanted responses may compete with elicitation of more desirable bnAbs. We currently have only a limited understanding of the diversity in the responses to SOSIP trimers that have been derived primarily from rabbit immunization studies (Bianchi et al., 2018; Klasse et al., 2018; McCoy et al., 2016; Pauthner et al., 2017). To investigate the specificity of polyclonal antibody responses, we recently developed an electron microscopy-based polyclonal epitope mapping (EMPEM) technique, whereby total serum IgG is isolated, digested into antigen-binding fragments (Fabs), and complexed with Env SOSIP.664 trimers before they are imaged using negative-stain or cryoelectron microscopy (cryo-EM). This technique provides a visual snapshot of the specificities present in polyclonal antibody preparations of an individual at a given time. We have thus far used EMPEM to define epitopes recognized by sera of BG505 SOSIP.664-immunized rabbits. The responses were shown to be quite similar between individual animals and to recognize a relatively narrow range of epitopes (Bianchi et al., 2018).
Here, we report EMPEM analysis of NHP polyclonal responses to BG505-based immunogens described in the Pauthner et al. (2017, 2019) studies. We found that high-titer animals recognized a greater diversity of epitopes than lower-titer animals. Our analyses also identified several strain-specific responses that might sterically block access to bnAb epitopes. Further, by using heterologous SOSIP trimers, we were able to map the epitope of cross-reactive antibodies that were detected in several immunized animals. It was found to be located at the gp120/gp41 interface region that includes the fusion peptide (FP). Finally, our ~3.9-Å resolution cryo-EM reconstruction of Fabs from a fully protected animal complexed to SOSIP trimers illustrated that one of the specificities, V1/V3, was likely a clonal response. This high-resolution structure also revealed that the V1/V3 loop antibody that resembles N332 supersite bnAbs at low resolution primarily interacts with the V1 loop, and that their binding would sterically compete with N332 supersite-specific bnAbs. In total, our imaging results provide many insights for Env trimer immunogen redesign including both positive and negative engineering opportunities.
RESULTS
Humoral Responses in Naive Macaques after SHIVBG505 Infection
The SHIVBG505 virus challenge study enrolled 12 immunized and 6 naive control animals (Pauthner et al., 2019). As previously described, the challenge virus contained a S375Y mutation that confers replication capacity in rhesus macaques and results in AIDS-like pathology (Pauthner et al., 2019). Animals received up to 12 intrarectal challenges at weekly intervals with a break after 6 challenges for those animals not infected early. Five of six naive animals became infected after the first challenge, and one after the second. In the low-titer NHP group, 2/6 animals became infected after the first challenge, and 4/6 after the second. Finally, in the high-titer NHP group, 1/6 animals became infected after the first challenge, 1/6 after the sixth, 1/6 after the tenth, 1/6 after the twelfth, and 2/6 never became infected (Pauthner et al., 2019).
Using EMPEM, we first analyzed the antibody responses in week 20 postchallenge (p.c.) sera (i.e., 20 weeks after first challenge) of the six non-immunized animals by forming complexes with BG505 SOSIP.664v5.2 Env glycoprotein trimers that were sequence matched to immunogen and the Env of the SHIVBG505 challenge virus. Figure 1 summarizes the EMPEM results wherein each dataset was reconstructed into a global 3D average and then subjected to subsequent 3D classification that sorted data into unique classes (Figure S1). Two of the six serum samples analyzed, BZ15 and BZ31, were not amenable to 3D reconstruction. The epitopes recognized in these animals, however, could be discerned from the 2D class averages when compared with known NHP monoclonal 2D class references (Figures S1D and S1E). One animal, CB00, showed no detectable binding to the BG505 trimer probe (Figure S1F), yet this was not unexpected because this animal showed waning BG505 S375Y pseudo-virus neutralizing titers 12 weeks p.c. (Pauthner et al., 2019), suggesting viral evolution away from the infecting virus and concomitant antibody evolution. The remaining three animals exhibited antibody responses in and around the N289/S241 glycan hole region (GH-1/GH-2) that is a highly immunogenic epitope in rabbits (McCoy et al., 2016; Bianchi et al., 2018; Klasse et al., 2018) and in the C3/V5 region of gp120 that we have previously described as the “C3/T465 epitope” in NHPs. Thus, considering a sample of only six animals and a time span of just 20 weeks under viral challenge pressure, the infected control macaques exhibited a relatively narrow repertoire of antibody epitope responses to the founding virus, with four different 3D Fab phenotypes detected against two regions of Env (Figures 1B and 1C). Notably, and not unexpectedly, virus challenges did not induce any antibodies that bound to the base of the trimer (Figure 1), which in rabbits is typically immunodominant on the soluble trimer and may limit elicitation of neutralizing immune responses (Bianchi et al., 2018; Cirelli et al., 2019; McCoy et al., 2016).
Figure 1. Polyclonal Antibody Responses of Naive Macaques Infected by SHIVBG505 Challenge.
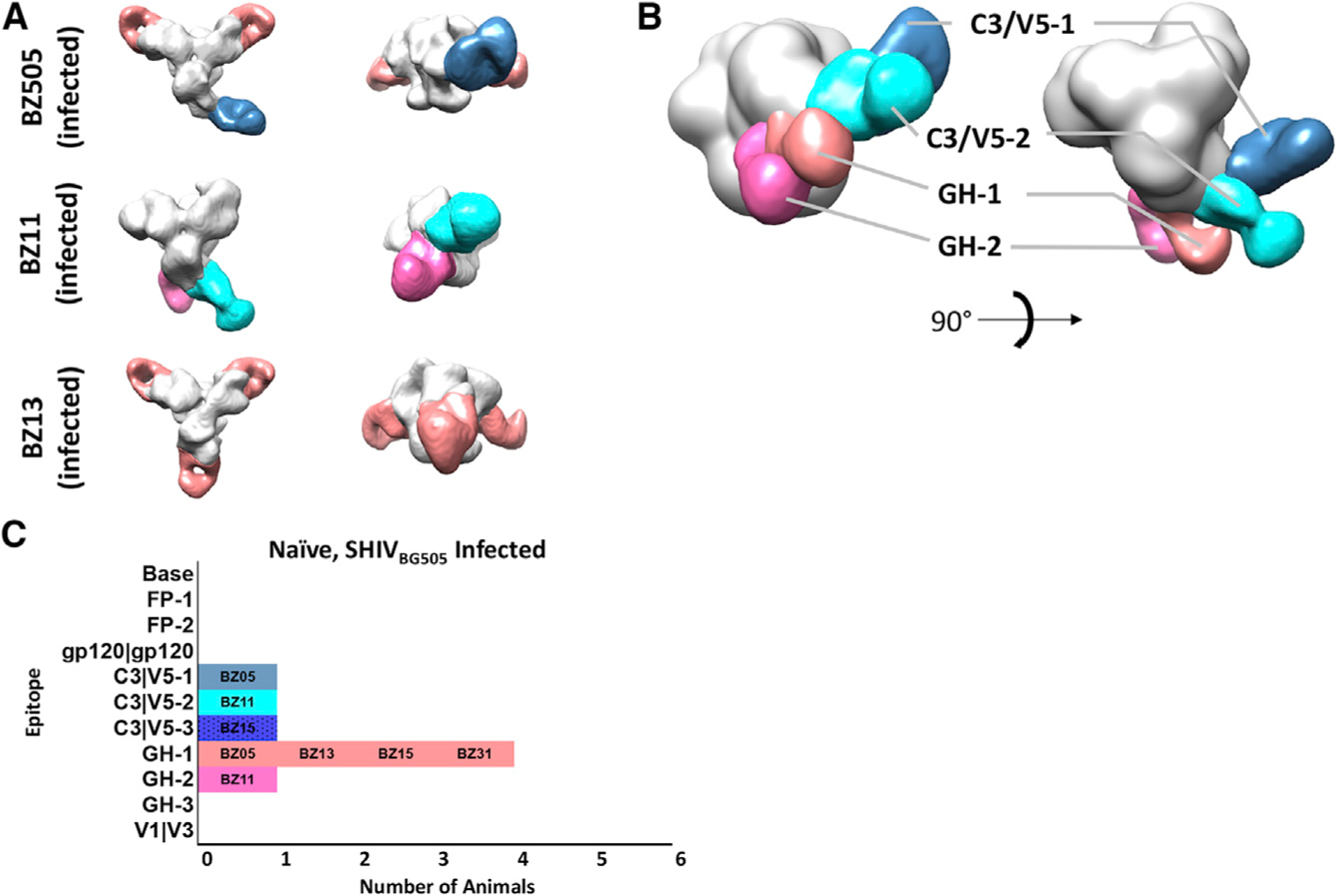
(A) Segmented 3D reconstructions of polyclonal Fab-BG505 trimer complexes from animals BZ05, BZ11, and BZ13.
(B) Composite 3D reconstruction demonstrating the antigenic map of epitopes amenable to 3D reconstruction observed in SHIVBG505-infected macaques (see Figure S1 for 3D classification and 2D class averages of animals BZ31, BZ15, and CB00).
(C) Bar graph summary of all the epitopes targeted by SHIVBG505-infected macaques (BZ15 phenotype represented in bar graph by 2D classification only).
Low-nAb-Titer-Vaccinated Animals Show Increased Diversity of Epitopes Compared with Infected Naive Controls
The next group that we examined were the low-titer animals subjected to a total of four identical immunizations with BG505 SOSIP.664 variants, including a booster injection 4 weeks prior to the first SHIVBG505 challenge. We performed EMPEM analysis on these animals 4 weeks after this last booster, immediately prior to the first virus challenge (i.e., week 68). These samples showed an overlapping set of epitopes but with increased diversity relative to the infected naive controls (Figures 1 and 2). The primary differences were that immunized animals also mounted an antibody response to the FP region, the gp120/gp120 inter face, and the base of the trimer. Despite the diversity of epitopes recognized, all animals from this low-titer group became infected within the first two virus challenges (Pauthner et al., 2019). Second to base binding, which is present in all BG505 SOSIP-immunized animals (McCoy et al., 2016; Bianchi et al., 2018), the most common epitopes targeted in the low-titer animals were the glycan hole 1 (GH-1) epitope, which was also the most frequently recognized epitope among the unimmunized controls, and the FP (Figures 1C and 2G). Thus, despite the presence of a diversity of vaccine-induced antibodies, this group of animals was not protected from the virus challenge. We note that none of the animals in this group had nAb titers above the ~1:500 threshold determined as the main correlate of protection (Pauthner et al., 2019).
Figure 2. Polyclonal Antibody Responses of Low Autologous Neutralizing Titer Macaques Immediately before SHIVBG505 Challenge.

(A–F) Segmented 3D reconstructions of (A) 12–060, (B) 12–065, (C) OQ7, (D) 12–153, (E) 12M-248, and (F) 12–149.
(G) Bar graph summary of the epitopes targeted by low autologous titer macaques. The number in parentheses indicates the number of SHIVBG505 challenges it took for the animal to become infected.
See Figure S2 for additional 3D phenotypes.
High-nAb-Titer-Vaccinated Animals Show the Most Diverse Humoral Responses
Relative to the infected naive and the low-titer-immunized macaques, the high-titer animals exhibited overlapping yet further diversified responses; their antibody response not only differed in the number and types of epitope recognized, but also in the apparent angles of approach for similar epitope clusters (Figures 1, 2, and 3). This higher diversity probably also gave rise to an increased Fab occupancy in the complexes observed in these animals. Figure 3 shows the results of the EMPEM analysis of week 68 serum samples from the high-titer macaque group that showed protection from infection and reduced peak viral loads relative to naive animals (Pauthner et al., 2017, 2019). Notably, two animals, 12–046 and 4O9, remained uninfected after 12 SHIVBG505 challenges (Pauthner et al., 2019).
Figure 3. Polyclonal Antibody Responses of High Autologous Neutralizing Titer Macaques Immediately before SHIVBG505 Challenge.

(A–F) Segmented 3D reconstructions of (A) 12–143, (B) 12–137, (C) 12M-169, (D) 11M-088, (E) 12–046, and (F) 4O9. The number in parentheses indicates the number of SHIVBG505 challenges it took for the animal to become infected (see Figure S3 for representative additional 3D phenotypes).
(G) Bar graph summary of the epitopes targeted by high autologous neutralizing titer macaques.
(H) Comparison of the angle of approach between bnAb PGT121 and animal 409 V1/V3 polyclonal response.
One striking difference between the low-titer- and high-titerimmunized animals was the appearance of antibodies binding to the V1/V3 region near the apex of the trimer in four out of six high-titer animals compared with none of the low-titer animals. At low resolution these responses resembled known bnAbs such as PGT121 that bind to the conserved glycine, aspartate, isoleucine, arginine (GDIR) co-receptor site and the N332 glycan (Figure 3H). Another notable phenotype for the high-titer animals was a different angle of approach by which Fabs bound to the C3/V5–3 epitope: three of six animals exhibited this phenotype, with the low-titer macaques favoring the GH-1 in lieu of this putative high-titer correlate (compare Figure 2 with Figures 3A, 3B, and 3F). This observation was consistent in additional unchallenged low- and high-titer animals from the immunization study (Figure S3D; Cirelli et al., 2019; Pauthner et al., 2017). Overall, the high-titer group displayed an increased diversity in the targeted epitopes, especially with the appearance of V1/V3 binding antibodies, and an associated increase in trimer occupancy (Figure 3), with apparently unique angles of approach as well, factors that may have contributed to these animals’ resistance to infection.
Evolution of Humoral Immune Responses
Taking advantage of the intensive schedule of blood draws throughout the immunization experiment, we applied EMPEM across multiple time points to study the evolution of antibody responses; we performed EMPEM analysis on available samples from high-titer animals after each booster immunization in an attempt to discern hierarchical signatures of developing protective antibody responses. The week 10 neutralization titers were 1:48, 1:83, and 1:404 for animals 12–137, 11M-088, and 4O9, respectively (Pauthner et al., 2017). Within this subset of animals there is no clear phenotype that correlated with future ability to protect from an autologous virus challenge. For example, both 12–137 and 4O9 displayed similar epitope recognition, namely, C3/V5–3 and FP, but 12–137 was infected after six intra-rectal challenges, whereas 4O9 remained uninfected after all 12 challenges. Serum samples from weeks 26–28 showed an increase in the diversity of epitopes recognized for animals 11M-088 and 4O9, which was not the case for 12–137. In fact, 12–137 and another member of the high-titer group that showed only marginal protection, 12–143, did not diversify the epitopes targeted after the first boost (see also Figure 3A).
Notably, fully protected animal 4O9 mounted V1/V3-specific antibodies as early as week 26 (post-boost 2), and these were not found in 11M-088, even after extensive 2D and 3D classification (Figure S4). Interestingly, we did not detect V1/V3-specific antibodies in 12–046, the other fully protected animal, at this stage, although this may have been a consequence of sample limitation and thus substantially lower Fab concentration during complex formation (see Method Details and Figure S4D). However, the weeks 66 and 68 (post-boost 3) 3D reconstructions demonstrate that both 11M-088 and 12–046 ultimately generated diverse antibody responses, with the V1/V3 loop now present in these animals’ polyclonal antibody responses and neutralization titers increased relative to week 26 (Figures 3E and 4B; Pauthner et al., 2017, 2019).
Figure 4. Polyclonal Antibody Responses of High Autologous Neutralizing Titer Macaques Showing the Evolution of Polyclonal Immune Responses among Animals that Exhibited Various Degrees of Protection to SHIVBG505 Challenge.

(A–C) Segmented 3D reconstructions of (A) 12–137, infected after the 6th SHIVBG505 challenge; 11M-088, infected after the 10th SHIVBG505 challenge (B); and 4O9, fully protected (C).
Top views (top) and side views (bottom) are shown for each time point.
See also Figure S4.
Thus, based on these EMPEM analyses, the ability to diversify the repertoire of epitopes recognized following continued booster immunizations generally coincided with higher neutralization titers and protection from infection.
Polyclonal Antibody Imaging Suggests Responses to the FP Are Responsible for Neutralization Breadth
Pauthner et al. (2017) reported sporadic neutralization breadth in a subset of the animals’ week 26 immunization sera using the 12-virus global pseudovirus panel, with the 398F1 virus strain being the most sensitive to neutralization (deCamp et al., 2014). Notably, this virus, like BG505, also exhibits a GH around position 289, which was shown to be very immunogenic and which was recognized by antibodies observed in 12–046 (Figure 3E), among others. To determine the cross-reactive specificity(ies), we prepared complexes of HIV398F1 SOSIP.664 envelope trimers with week 25 or 26/28 Fabs from animals 12–137, 12–046, and 12–084 (this animal exhibited exceptionally high titers but was not among the challenge group) (Pauthner et al., 2017) and subjected them to EMPEM analysis (Figure 5A). Complexes with HIV25710 Env trimers and animal 12–046 week 26 serum were also examined (Figure S5B). These macaques had HIV398F1 neutralization titers of 1:32,620, 1:256, and 1:399, respectively, with 12–046 titers against 25710 at a more modest 1:160. Excluding antibodies to the trimer base, we observed only a single epitope specificity binding to heterologous trimers, namely, the FP region, being contacted at two slightly different angles of approach (Figures 5A and 5B; Figure S5B). These phenotypes are reminiscent of bnAbs PGT151, VRC34, and ACS202, all of which include the FP as part of their epitope (Figure 5C; Kong et al., 2016; Lee et al., 2016; Yuan et al., 2019). Interestingly, the FP sequence is identical between BG505, 398F1, and 25710 (Figure S5A). Thus, based on these EMPEM analyses of a limited subset of heterologous viruses that were sensitive to neutralization by immunized macaque polyclonal sera, it appears that antibodies to the FP may be one of the main sites responsible for the breadth observed in several animals (Pauthner et al., 2017).
Figure 5. Polyclonal Sera Cross-Reactivity Mapping.

(A) SOSIP version of virus 398F1 Env trimer complexed with high-titer animals 12–084, 12–137, and 12–046.
(B) Summary of epitopes targeted on 398F1 by high-titer animals.
(C) Comparison of polyclonal FP approach angles with bnAbs PGT151 and VRC34..
NT, neutralization titer.
See also Figure S5.
3D Cryo-EM Reconstruction Reveals Structural Details of the Macaque “V1/V3” and “C3/V5” Targeting Antibodies
Given the similarity of the apical antibodies that we detected in 4/6 high-titer animals to N332 bnAbs, we attempted further characterization of this specificity by conducting high-resolution cryo-EM analysis. We used week 26 serum from the fully protected animal 4O9 (Figures 3F and 4C) to make complexes with BG505 SOSIPv5.2 to assess the structural details of the epitope recognition in high-titer animals (summarized in Figure 6). The data produced a ~3.9-Å global resolution 3D reconstruction (Figure 7A). Notably, relative to our more typical cryo-EM sample preparation conditions using monoclonal antibodies (mAbs), we prepared the polyclonal cryo-EM sample using slightly modified conditions to maximize the likelihood of observing as many of the epitopes as possible (see Discussion and STAR Methods). Consequently, by attempting to maximize the Fab:trimer ratio, we generated complexes in which the Env trimers were saturated with Fabs, and we were not able to classify out trimers occupied by less than the 9 Fabs depicted in Figure 7.
Figure 6. Summary of Epitopes Targeted by NHPs.

(A) Venn diagram showing shared phenotypes among pre-challenge low- and high-titer animals and non-immunized infected control animals.
(B) Combined global epitope summary of antibodies observed for low-titer, high-titer, and infected unimmunized control animals.
See also Figure S6.
Figure 7. Epitopes Targeted by Fully Protected Animal 409.

(A) Low (left) and high (right) contour cryo-EM resolution maps showing that the epitope-paratope regions of the V1 interactions are resolved at <4.0Å, with the C3/V5 and base responses showing lower resolutions.
(B and C) Map and model of V1/V3 antibody with docked Fab demonstrating how the antibody inserts between glycans at positions 133 and 137. In (C), the map has been removed for clarity.
(D) Close-up view of V1 interaction showing proximity to and potential blocking of GDIR interaction.
(E) Overlay of BG505 SOSIP.664 GDIR-V1 loop configurations of ground state (in the absence of an N332 bnAb) (PDB: 5V8M) and with N332 glycan supersite bnAbs PGT128 and PGT121 (PDB: 5ACO and 5CEZ, respectively) bound. The V1 loop in the animal 409 polyclonal complex is in the ground state, which likely precludes access to the underlying GDIR motif and therefore unlikely to develop breadth.
(F) Negative-stain reconstructions of monoclonal antibodies P1L4, P2C23, P2G23, and P2L14 isolated from animal 409. All antibodies (shown in blue) target the C3/V5 epitope similar to what we observe in the week 64 polyclonal response.
See also Figure S7.
In the global 3D reconstruction, we observed clear density for Fabs corresponding to the trimer base, V1/V3, and C3/V5–3 epitopes. The density for the trimer base Fabs was diffuse and poorly resolved, suggesting a heterogenous mixture of base antibodies were being averaged together. Conversely, the V1/V3 and, to a lesser extent, the C3/V5–3 Fab densities were remarkably well resolved (Figure 7A). These data enabled more in-depth analyses of the molecular interactions of these two paratopeepitope interfaces. The V1/V3 Fabs were so well resolved that they likely corresponded to a single clonal lineage and enabled us to determine the CDR loop lengths to be ~10, ~14, and ~5 residues long for CDRH3, CDRH2, and CDRH1, respectively (Kunik et al., 2012). Further, the less variable regions of the Fab were resolved to sufficient resolution to semi-confidently model side chains.
The primary interaction between the V1/V3 Fab and the apex of the trimer is between the CDRH3 of the Fab, inserted between glycans at positions N133 and N137 to make peptide contacts at residues N137 and I138, T139 of the trimer V1 loop, as well as the CDRL3, which contacts R143 within the same region (Figures 7B–7D). We previously reported virus neutralization data that implicated an epitope that includes residues N133 and N137, which are part of the V1 loop, as part of these antibodies binding (Klasse et al., 2018). Further, Pauthner et al. (2017) reported up to nearly 100-fold decreases in neutralization titer in this animal upon amino acid insertions in the vicinity of these residues. Finally, the CDRH1 loop makes side-chain contacts with R327 (Figure S7H), which is a residue in the GDIR motif of the co-receptor binding site.
The V1 loop of Env in the 4O9 Fab-complexed structure is in a nearly identical conformation relative to the uncomplexed BG505 SOSIP.664 trimer control, which we deem the ground-state conformation that effectively masks the conserved GDIR co-receptor binding site (Sok et al., 2016). Conversely, both PGT121 and PGT128 bnAbs binding displaces the V1 from this conformation for efficient recognition of N332 and the GDIR motif (Figure 7E; Lee et al., 2015). The cryo-EM map shows that although the main interaction is between the V1 loop and the CDRH3 of the Fabs, some contacts between the V3 loop and the CDRH1 and CDRH3 of the Fabs are present, with the N332 clearly pushed out of the way (Figure S7G). This type of interaction is also observed in a rabbit nAb recently described (Nogal et al., 2019). Given the high sequence and glycosylation variability in V1 among global HIV isolates, this epitope is likely strain specific, with limited potential for broadening. The substantial epitope overlap of the NHP V1 antibodies with the N332 supersite may constitute a roadblock for the development of bnAbs to this region.
Although the less well-resolved C3/V5 epitope precluded contact residue assignment, we discerned that the primary interaction is between the BG505 variable loop 5 and the light chain of the antibody, with the glycan at position 465 shifted out of the way to accommodate the contacts (Figure S7B). The CDRH3 appears to be relatively short, possibly in order to accommodate a glycan at position 355 (Figures S7B and S7C). Notably, negative stain reconstructions of monoclonal nAbs isolated from this same animal also bound to this site (Figure 7F), with the same angle of approach as in both the cryo-EM and negative-stain reconstructions of weeks 10, 26, and 64 (Figures 4C and 5A). These nAbs only partially account for the serum neutralization, further emphasizing the potential role of other epitope specificities, namely, the V1 response described above.
DISCUSSION
In an attempt to map NHP antibody responses in the context of both differing neutralization titers and levels of protection from a SHIV challenge (Pauthner et al., 2017, 2019), we applied our EMPEM approach to identify and characterize vaccine-induced antibodies that correlated with serum neutralizing titers and, consequently, protection (Pauthner et al., 2019). In contrast with rabbits, where the 241/289 GH and trimer base are the predominant targets of the antibody response, several additional epitopes were identified to be immunogenic in NHPs, some of which are likely responsible for the bulk of the neutralization activity (Cirelli et al., 2019; Klasse et al., 2018; Pauthner et al., 2017). These epitopes include the FP, V1/V3, and a newly identified epitope at the gp120/gp120 interface.
Antibodies elicited following SHIVBG505 infection of unimmunized animals revealed a relatively focused antibody response to a series of epitopes stretching from the 241/289 GH to the C3/V5 region and comprised only a subset of those observed in the immunized animals. We hypothesize that one potential reason for the differences between infection and vaccination may be due to the persistent antigen exposure and viral escape in the SHIV-infected group that drives continuous evolution of focused immune responses to the two immunodominant regions on BG505 viral Env, namely, the 241/289 GH and the C3/V5 super-epitope. In other words, viral infection maintains focused germinal center responses, whereas prime-boost subunit vaccination results in more diverse epitope responses that may be influenced by the presence of circulating antibodies. We cannot, however, rule out that the early and immunodominant presence of base binding antibodies in the vaccine group may assist antigen recruitment to secondary lymphoid organs and germinal centers, and/or may result in immune complexes that present the envelope trimers in a unique orientation to B cells.
In immunized animals, high neutralization titers coincided with a more diverse epitope recognition compared with low-titer animals. Aside from one animal (12–046), this increased epitope diversity of the responses observed appeared to be associated with neutralization and protection. Neutralization was associated with epitope specificities that differed from those of the much more focused nAb response observed in immunized rabbits (Bianchi et al., 2018). However, an increased diversity of epitopes recognized alone did not automatically result in improved protection from infection. For example, animal 12M-248, which exhibited the highest diversity of epitopes in the low-titer group, did not exhibit an increased neutralization or protection (Figure 2; Pauthner et al., 2019), whereas animal 12–046 was completely protected despite a rather limited set of recognized epitopes. Also, animal 11M-088 mounted antibodies to two more epitopes relative to the fully protected animals and one more relative to 12M-169, yet this animal was infected the earliest of all the animals that targeted the V1/V3 epitope (10 challenges). Interestingly, two of the four animals in the low-titer group that became infected after only two challenges had more diverse responses than those that became infected after a single challenge (Figure 2E). Thus, although diversification of recognized epitopes may increase the likelihood of neutralization or protection, the overall titers of nAbs appeared to be primarily responsible for protection.
C3/V5 was highly immunogenic, and we found a range of approach angles for antibodies binding to this epitope. Some, namely, those designated C3/V5–3 antibodies, were detected only in high-titer animals from the vaccination study (Figure 3), a finding that is consistent with a prior study (Cirelli et al., 2018). Still, without the additional presence of V1/V3 antibodies, the C3/V5–3 epitope by itself was insufficient for durable protection in the face of repetitive SHIV challenges. Indeed, viruses isolated from animal 12–137, which became infected after six challenges, displayed putative escape mutations at positions 354 and 356 (Pauthner et al., 2019). Because these residues flank the N355 glycan, which is part of the C3/V5–3 epitope (Pauthner et al., 2019), it appears that either the escape threshold for this epitope is relatively low, or that antibodies against this epitope exert a strong selective pressure for escape. Also, given its overlapping footprint with the CD4bs, C3/V5–3 class antibodies will compete with and potentially suppress the induction of CD4bs bnAbs such as VRC01.
Despite being a potential correlate of protection in this NHP challenge model, and closely resembling the bnAb PGT121 in epitopic footprint (Figure 3H), our cryo-EM analysis reveals that the V1/V3 class of antibodies is likely competitive and counterproductive to the induction of N332 supersite bnAbs (Figure 7D). Thus, low-resolution EMPEM can be used to down-select which complexes are worth pursuing at high resolution to elucidate the fine details of epitope-paratope interactions and compare with bined bnAbs. However, low-resolution EMPEM may also be com-with traditional serological assays such as ELISAs and mutational mapping with pseudovirus-based neutralization assays to generate similar observations and dissect the molecular differences between strain-specific and broad neutralizing responses. Should the human humoral response to the V1/V3 be analogous to rhesus macaques, our high-resolution structure suggests that additional engineering may be necessary to optimize the N332/GDIR epitope presentation in this immunogen.
Although detection of FP-like antibodies in rabbits was rare (Bianchi et al., 2018), FP responses were readily detectable in rhesus macaques as early as week 10. We also show that FP epitope likely contributed to the observed neutralization breadth in some animals and is therefore the most promising antibody response elicited by the BG505 trimer immunization. Further, because the FP is a well-characterized target of human bnAbs, our results suggest that macaques are a suitable preclinical model for evaluating FP-directed immunogens intended to elicit PGT151-, ACS202-, or VRC34-like bnAb.
With the first dose-escalation phase I clinical trial using the BG505 SOSIP.664 gp140 trimer as the immunogen now recruiting human subjects (ClinicalTrials.gov: NCT03699241), the present findings are an important preclinical benchmark. We show that, similar to rabbits, the macaque preclinical model develops polyclonal responses in a tiered fashion, with antibodies against the most accessible immunogenic epitopes (i.e., the trimer base) elicited early, followed by more diverse and individualized responses. We predict that human responses will be similar to NHPs, and our EMPEM approach will enable rapid comparison with this end. Moreover, the present structural information can be exploited to design epitope knockout trimer probes that can be used to quantify epitope-specific B cell responses by fluorescence-activated cell sorting (FACS) or that can be used for the isolation of B cell subsets specific for individual epitope(s); a more detailed characterization of mAbs will offer further lessons for immunogen redesign. Finally, our detailed structural analyses identified potential weak points of the current generation of BG505 SOSIP trimer immunogens and will help to inform the design and engineering of future iterations of this immunogen.
STAR★METHODS
LEAD CONTACT AND MATERIALS AVAILABILITY
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Andrew B. Ward (andrew@scripps.edu). This study did not generate new unique reagents.
EXPERIMENTAL MODEL AND SUBJECT DETAILS
Rhesus Macaques
The plasma and serum samples used in this work derive from the previously described rhesus macaque immunization and subsequent challenge experiments using a subset of the cohort (Pauthner et al., 2017, 2019). Briefly, 70 rhesus macaques (3–4 years old at the time of first immunization) received a prime and 2 booster immunizations over the course of 26 weeks. Of this immunized cohort, 6 high and 6 low neutralization titer animals were selected to enter the challenge study, with 6 non-immunized (“naïve”) animals used as controls. The sexes of the animals as reported in Pauthner et al., 2018 are as follows: 11 Females (12–137, 12–143, 12M-169, 11M-088, 4O9, 12–153, 12–065, 12–149, 12M-248, 0Q7, BZ13), 7 Males (12–046, 12–060, BZ05, BZ11, BZ15, BZ31, CB00). The Scripps Research Institutional Animal Care and Use Committee (IACUC) approved all experimental procedures involving all the animals.
METHOD DETAILS
Soluble Env Protein Production
BG505 SOSIP.664 v5.2, 25710, X1632, CE1176, and 398F1 SOSIP.664 were co-transfected with furin into HEK293F cells, followed by affinity chromatography using either 2G12 or PGT135 as the ligands. Affinity eluents were then purified via size exclusion chromatography (SEC) in order to remove trimer aggregates or protomers. Full details of trimer purification can be found elsewhere (Pugach et al., 2015).
Plasma or Serum IgG Purification
Serum or plasma polyclonal IgGs were Protein A affinity-purified using a 1:1 ratio of resin to undiluted serum or plasma sample. After a 4-to-5-fold dilution with PBS, samples were incubated at room temperature (RT) for 5 h or overnight at 4°C, followed by a 3-fold wash step using 10 volumes of PBS. Elution consisted of 5–10 volumes of 0.1M glycine, pH 2.5 into vessels pre-conditioned with ~20% v/v Tris-HCL, pH 8. Eluents were buffer exchanged into TBS by centrifugation with 10 kDa cutoff membranes (EMD Millipore).
IgG Digestion
In order to perform EM imaging, Fabs were produced by digesting the IgGs using resin-immobilized papain (50 μL settled resin/mg IgG, Thermo fisher Scientific). The digestion occurred in 20 mM sodium phosphate, 10 mM EDTA, 20 mM cysteine, pH 7.4 for 5–6 h at 37°C. In order to remove Fc and intact IgG, the digestion mix was incubated with protein A Sepharose (GE Healthcare) for 1 h at RT. The Fab-containing supernatant was buffer exchanged into TBS by centrifugation with 10kDa cutoff membranes (EMD Millipore).
EM Complex Preparation
Complexes for negative-stain EM were prepared similarly to the previously described method, except ELISA EC50 was not used to guide serum Fab concentration to be used (Bianchi et al., 2018). Briefly, after buffer exchanging into TBS, up to ~1 mg of total Fab was incubated overnight with 10 μg BG505 trimers at RT in ~50 μL total volume. Complexes were then purified by SEC using Superose 6 Increase 10/300 column (GE Healthcare) in order to remove unbound Fab. The flow-through fractions containing the complexes were pooled and concentrated using 100 kDa cutoff centrifugal filters (EMD Millipore). The final trimer concentration was titrated to ~0.04 mg/mL prior to application onto carbon-coated copper grids. Of note, depending on sample availability, not all complexes were prepared using identical Fab concentrations during incubation with trimers, with samples from animals where serum material was limiting resulting in lower Fab:trimer ratio. For animal 12–046, week 10 and 26 samples were insufficient to include in the time-course study as the purified total Fab concentration was severalfold lower than the rest of the animals (see Table S1).
Negative-Stain EM
The SEC-purified complexes were applied to glow-discharged, carbon-coated 400-mesh copper grids, followed by pipetting 3 μL of 2% (w/v) uranyl formate stain and blotting, followed by application of another 3 μL of stain for 45–60 s, again followed by blotting. Stained grids were stored under ambient conditions until ready for imaging. Images were collected via Leginon software using a Tecnai T12 electron microscopes operated at 120 kV and 52,000× magnification (Suloway et al., 2005). In all cases, the electron dose was 25 e−/Å 2. Particles were picked from the raw images using DoG Picker and placed into stacks using Appion software (Lander et al., 2009). 2D reference-free alignment was performed using iterative MSA/MRA. The particle stacks were then converted from IMAGIC to RELION-formatted MRC stacks and subjected to RELION 2.1 2D and 3D classification (Scheres, 2012).
Cryo-EM
Animal 4O9 complex preparation for cryo-EM was modified relative to the negative stain workflow in order to maximize trimer occupancy while still resulting in sufficient complex concentration for cryo-EM grids. Here, ~50 μg of trimer was incubated with > 3 mg of Fab and incubated overnight as indicated above. The complex was then concentrated to 1.5 mg/mL and mixed with Lauryl Maltose Neopentyl Glycol (LMNG, Anatrace) prior to deposition onto 2/2 Quantifoil grids (EMS) that were glow discharged for 10 s directly preceding the deposition in a Vitrobot (Thermo Scientific). Once sample was deposited the grids were blotted and plunged into liquid ethane using the Vitrobot to immobilize the particles in vitreous ice. Using Leginon image acquisition software we collected 3,347 micrographs at a nominal magnification of 29,000x with a Gatan K2 summit detector mounted on a Titan Krios set to 300 kV set to counting mode for the data collection (Suloway et al., 2005). The dose rate was ~4.78 e-/pix/s with frame exposure of 250 ms, with a total exposure time and dose of 13.5 s and 61 e−/Å2, respectively. MotionCor2 was used for frame alignment and CTF models were determined using GCTF (Zheng et al., 2017). DogPicker was used to pick 666,007 particles, which were then extracted and subsequently 2D-classified in cryoSPARC (Punjani et al., 2017; Voss et al., 2009). Selected 2D classes amounting to 277,453 particles were then fed into the 3D heterogeneous refinement algorithm with 3 classes requested. Of these, only one 3D class (155,394 particles, GSFSC 6.21Å) amounted to a trimer with Fabs bound and it was advanced to homogeneous refinement using C3 symmetry, which resulted in a GSFSC resolution of 4.01Å. Subsequent non-uniform C3 symmetry refinement resulted in a final GSFSC resolution of 3.87Å (see Table S2 for data collection parameters).
Model Building and Refinement
The BG505 SOSIP.664 trimer structure from PDB 5ACO was docked into the cryo-EM map using UCSF Chimera (Pettersen et al., 2004). Subsequent iterations of manual and Rosetta fragment library based centroid rebuilding and refinement were then performed (Simoncini et al., 2012).The resulting model was then refined using all atom refinement under constraints of the density map. Glycans were manually built in Coot (Emsley et al., 2010).
Isolation of Monoclonal Antibodies
Single cell sorting was performed as previously described (Mason et al., 2016). In brief, rhesus PBMCs were stained with an antibody cocktail of CD3 (clone SP34–2, BD Biosciences), CD4 (clone OKT4, BioLegend), CD8 (clone RPA-T8, BD Biosciences), CD14 (M5E2, BD Biosciences), CD20 (clone 2H7, Biolegend), IgM (clone MHM-88, Biolegend), IgG (clone G18–145, BD Biosciences) and BG505 SOSIP-AviB conjugated with streptavidin-Alexa Fluor 647 (AF647) and streptavidin-Alexa Fluor 488 (AF488) respectively. Cells were stained at room temperature for 20 mins in the dark. Antigen-specific memory B cells (CD3−CD4−CD8−CD14−CD20+IgM−IgG+BG505 SOSIP.664 dual positive) were single-cell sorted into 384-well culture plates according to the published protocol (Huang et al., 2013). After 2 weeks of culture, supernatants were harvested and screened for neutralization activity. Ig genes from neutralization positive wells were amplified by RT-PCR, nested PCR, then cloned into expression vectors and expressed in 293F cells.
QUANTIFICATION AND STATISTICAL ANALYSIS
Due to the small animal subject sample sizes and thus insufficient power to detect significant polyclonal response differences among the unimmunized naive, low titer, and high titer animals, no statistical tests were applied for the purpose of describing the trends in responses and their diversity. Furthermore, as the serum samples were themselves sometimes a limiting factor, accurate quantitation and subsequent quantitative comparison among individual animals was not possible.
DATA AND CODE AVAILABILITY
3D EM reconstructions have been deposited in the Electron Microscopy Databank (http://www.emdataresource.org/) or Protein Databank (http://www.rcsb.org) under the accession numbers listed in the Key Resources Table.
KEY RESOURCES TABLE
| REAGENT OR RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Biological Samples | ||
| Plasma or serum from rhesus macaques | (Pauthner et al., 2017, Pauthner et al., 2018) | N/A |
| Chemicals, Peptides, and Recombinant Proteins | ||
| BG505 SOSIP.664 | Produced in house (Sanders et al., 2013) | N/A |
| 398F1 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| 25710 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| CE1176 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| X1632 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| Protein A Resin | GE Healthcare | Cat# 17-5280-02 |
| Superose 6 Increase Size Exclusion Column | GE Healthcare | Cat# 17517201 |
| Uranyl Formate | Electron Microscopy Sciences | Cat# D310 25 GM |
| Lauryl Maltose Neopentyl Glycol | Anatrace | Cat# NG310 5 GM |
| Critical Commercial Assays | ||
| Pierce Fab Preparation Kit | Thermo Fisher Scientific | Cat# 44985 |
| Negative stain EM Grids | Electron Microscopy Sciences | Cat# EMS400-CU |
| Cryo-EM grids | Electron Microscopy Sciences | Cat# Q4100CR2 |
| Deposited Data | ||
| Cryo-EM Reconstruction of Polyclonal Serum Fab from a Protected Rhesus Macaque 4O9 Complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20396 |
| Negative-stain EM map of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20399 |
| Negative-stain EM map of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 | EMDataBank | EMD: 20400 |
| Negative-stain EM map of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20401 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–060 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20402 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12–065 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20403 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque OQ7 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20404 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12–153 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20405 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12M-248 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20409 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12–149 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20410 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–143 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20415 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20423 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12M-169 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20424 |
| Negative-stain EM map of BG505-immunized rhesus macaque 11M-088 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20425 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20426 |
| Negative-stain EM map of BG505-immunized rhesus macaque 4O9 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20427 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20428 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20946 |
| Negative-stain EM map of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20429 |
| Negative-stain EM map of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20970 |
| Negative-stain EM map of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20971 |
| Negative-stain EM map of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20972 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–084 wk 26 serum fab complexed with 398F1 SOSIP.664 | EMDataBank | EMD: 20992 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 26 serum fab complexed with 398F1 SOSIP.664 | EMDataBank | EMD: 20936 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–046 wk 26 serum fab complexed with 398F1 SOSIP.664 | EMDataBank | EMD: 20937 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20973 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20974 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20975 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20976 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20977 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20978 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20979 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20980 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20981 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–060 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20982 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–060 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20983 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–065 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20938 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–065 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20984 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–065 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20985 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque OQ7 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20939 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque OQ7 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20940 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–143 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20941 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–143 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20987 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20988 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20989 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20942 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20921 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20943 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20944 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20899 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20898 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20897 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20896 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20896 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20895 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20894 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20893 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20990 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20892 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20891 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20890 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20889 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20889 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20888 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20887 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20991 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20886 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–046 wk 26 serum fab complexed with 25710 | EMDataBank | EMD: 20885 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–084 wk 28 serum fab complexed with BG505 SOSIP trimer | EMDataBank | EMD: 20884 |
| PDB Coordinates of BG505 SOSIP.664 trimer complexed with Polyclonal Serum Fab from a Protected Rhesus Macaque 4O9 | Protein Data Bank | PDB ID: 6V0R |
| Experimental Models: Cell Lines | ||
| Human: FreeStyle HEK293F | Thermo Fisher Scientific | Cat# R79007 |
| Human: HEK294T | ATCC | Cat# CRL-3216 |
| Software and Algorithms | ||
| Unicorn 7.0 | GE Healthcare | https://www.gelifesciences.com/ |
| UCSF Chimera | Pettersen et al., 2004 | N/A |
| Appion database | Lander et al., 2009 | N/A |
| Leginon | Suloway et al., 2005 | N/A |
| DoG Picker | Voss et al., 2009 | N/A |
| Relion | Scheres, 2012 | N/A |
| CryoSparc | Punjani et al., 2017 | N/A |
Supplementary Material
Highlights.
Electron microscopy polyclonal epitope mapping of immunized and challenged macaques
Diversity of epitopes and specific angles of approach are hallmarks of protection
Neutralization breadth is due in part to fusion peptide-specific antibodies
Cryo-EM analysis of a protected animal details a putatively neutralizing paratope
ACKNOWLEDGMENTS
We would like to thank Bill Anderson and Hannah Turner for assistance with microscopy, Charles Bowman and J.C. Ducom for computational support, and Lauren Holden and Aleksandar Antanasijevic for critical reading of the manuscript. This work was supported by NIH grants R01 AI136621 and UM1 AI100663 (to A.B.W. and L.H.) and Swiss National Science Foundation fellowships P300PB_160969 and P3P3PB_160970 (M.B., Switzerland). C.A.C. is supported by a NIH F31 Ruth L. Kirschstein Predoctoral Award Al131873 and by the Achievement Rewards for College Scientists Foundation.
Footnotes
SUPPLEMENTAL INFORMATION
Supplemental Information can be found online at https://doi.org/10.1016/j.celrep.2020.02.061.
DECLARATION OF INTERESTS
The authors declare no competing interests.
REFERENCES
- Ahmed Y, Tian M, and Gao Y (2017). Development of an anti-HIV vaccine eliciting broadly neutralizing antibodies. AIDS Res. Ther 14, 50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianchi M, Turner HL, Nogal B, Cottrell CA, Oyen D, Pauthner M, Bastidas R, Nedellec R, McCoy LE, Wilson IA, et al. (2018). Electron-Microscopy-Based Epitope Mapping Defines Specificities of Polyclonal Antibodies Elicited during HIV-1 BG505 Envelope Trimer Immunization. Immunity 49, 288–300.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burton DR, and Hangartner L (2016). Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu. Rev. Immunol 34, 635–659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang G-Y, Geng H, Pancera M, Xu K, Cheng C, Acharya P, Chambers M, Druz A, Tsybovsky Y, Wanninger TG, et al. (2017). Structure-Based Design of a Soluble Prefusion-Closed HIV-1 Env Trimer with Reduced CD4 Affinity and Improved Immunogenicity. J. Virol 91, e02268–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cirelli KM, Carnathan DG, Nogal B, Martin JT, Rodriguez OL, Upadhyay AA, Enemuo CA, Gebru EH, Choe Y, Viviano F, et al. (2019). Slow Delivery Immunization Enhances HIV Neutralizing Antibody and Germinal Center Responses via Modulation of Immunodominance. Cell 177, 1153–1171.e28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen KW, and Frahm N (2017). Current views on the potential for development of a HIV vaccine. Expert Opin. Biol. Ther 17, 295–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen MS, Chen YQ, McCauley M, Gamble T, Hosseinipour MC, Kumarasamy N, Hakim JG, Kumwenda J, Grinsztejn B, Pilotto JHS, et al. ; HPTN 052 Study Team (2011). Prevention of HIV-1 infection with early antiretroviral therapy. N. Engl. J. Med 365, 493–505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- deCamp A, Hraber P, Bailer RT, Seaman MS, Ochsenbauer C, Kappes J, Gottardo R, Edlefsen P, Self S, Tang H, et al. (2014). Global panel of HIV-1 Env reference strains for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol 88, 2489–2507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emsley P, Lohkamp B, Scott WG, and Cowtan K (2010). Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr 66, 486–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Escolano A, Steichen JM, Dosenovic P, Kulp DW, Golijanin J, Sok D, Freund NT, Gitlin AD, Oliveira T, Araki T, et al. (2016). Sequential Immunization Elicits Broadly Neutralizing Anti-HIV-1 Antibodies in Ig Knockin Mice. Cell 166, 1445–1458.e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes BF, Shaw GM, Korber B, Kelsoe G, Sodroski J, Hahn BH, Borrow P, and McMichael AJ (2016). HIV-Host Interactions: Implications for Vaccine Design. Cell Host Microbe 19, 292–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu DC, and O’Connell RJ (2017). Progress in HIV vaccine development. Hum. Vaccin. Immunother 13, 1018–1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang J, Doria-Rose NA, Longo NS, Laub L, Lin C-L, Turk E, Kang BH, Migueles SA, Bailer RT, Mascola JR, and Connors M (2013). Isolation of human monoclonal antibodies from peripheral blood B cells. Nat. Protoc 8, 1907–1915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jardine J, Julien JP, Menis S, Ota T, Kalyuzhniy O, McGuire A, Sok D, Huang PS, MacPherson S, Jones M, et al. (2013). Rational HIV immunogen design to target specific germline B cell receptors. Science 340, 711–716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klasse PJ, Ketas TJ, Cottrell CA, Ozorowski G, Debnath G, Camara D, Francomano E, Pugach P, Ringe RP, LaBranche CC, et al. (2018). Epitopes for neutralizing antibodies induced by HIV-1 envelope glycoprotein BG505 SOSIP trimers in rabbits and macaques. PLoS Pathog. 14, e1006913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong R, Xu K, Zhou T, Acharya P, Lemmin T, Liu K, Ozorowski G, Soto C, Taft JD, Bailer RT, et al. (2016). Fusion peptide of HIV-1 as a site of vulnerability to neutralizing antibody. Science 352, 828–833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunik V, Ashkenazi S, and Ofran Y (2012). Paratome: an online tool for systematic identification of antigen-binding regions in antibodies based on sequence or structure. Nucleic Acids Res. 40, W521–W524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lander GC, Stagg SM, Voss NR, Cheng A, Fellmann D, Pulokas J, Yoshioka C, Irving C, Mulder A, Lau P-W, et al. (2009). Appion: an integrated, database-driven pipeline to facilitate EM image processing. J. Struct. Biol 166, 95–102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JH, de Val N, Lyumkis D, and Ward AB (2015). Model Building and Refinement of a Natively Glycosylated HIV-1 Env Protein by High-Resolution Cryoelectron Microscopy. Structure 23, 1943–1951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JH, Ozorowski G, and Ward AB (2016). Cryo-EM structure of a native, fully glycosylated, cleaved HIV-1 envelope trimer. Science 351, 1043–1048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu C-L, Murakowski DK, Bournazos S, Schoofs T, Sarkar D, Halper-Stromberg A, Horwitz JA, Nogueira L, Golijanin J, Gazumyan A, et al. (2016). Enhanced clearance of HIV-1-infected cells by broadly neutralizing antibodies against HIV-1 in vivo. Science 352, 1001–1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason RD, Welles HC, Adams C, Chakrabarti BK, Gorman J, Zhou T, Nguyen R, O’Dell S, Lusvarghi S, Bewley CA, et al. (2016). Targeted Isolation of Antibodies Directed against Major Sites of SIV Env Vulnerability. PLoS Pathog. 12, e1005537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCoy LE, van Gils MJ, Ozorowski G, Messmer T, Briney B, Voss JE, Kulp DW, Macauley MS, Sok D, Pauthner M, et al. (2016). Holes in the glycan shield of the native HIV envelope are a target of trimer-elicited neutralizing antibodies. Cell Rep. 16, 2327–2338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nogal B, McCoy LE, van Gils MJ, Cottrell CA, Voss JE, Andrabi R, Pauthner MG, Liang C-H, Messmer T, Nedellec R, et al. (2019). HIV Envelope Trimer-Elicited Autologous Neutralizing Antibodies Bind a Region Overlapping the N332 Glycan Supersite. bioRxiv. 10.1101/831008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauthner M, Havenar-Daughton C, Sok D, Nkolola JP, Bastidas R, Boopathy AV, Carnathan DG, Chandrashekar A, Cirelli KM, Cottrell CA, et al. (2017). Elicitation of Robust Tier 2 Neutralizing Antibody Responses in Nonhuman Primates by HIV Envelope Trimer Immunization Using Optimized Approaches. Immunity 46, 1073–1088.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauthner MG, Nkolola JP, Havenar-Daughton C, Murrell B, Reiss SM, Bastidas R, Prévost J, Nedellec R, von Bredow B, Abbink P, et al. (2019). Vaccine-Induced Protection from Homologous Tier 2 SHIV Challenge in Nonhuman Primates Depends on Serum-Neutralizing Antibody Titers. Immunity 50, 241–252.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, and Ferrin TE (2004). UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem 25, 1605–1612. [DOI] [PubMed] [Google Scholar]
- Prabakaran P, Chen W, and Dimitrov DS (2014). The Antibody Germline/Maturation Hypothesis, Elicitation of Broadly Neutralizing Antibodies Against HIV-1 and Cord Blood IgM Repertoires. Front. Immunol 5, 398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pugach P, Ozorowski G, Cupo A, Ringe R, Yasmeen A, de Val N, Derking R, Kim HJ, Korzun J, Golabek M, et al. (2015). A native-like SOSIP.664 trimer based on an HIV-1 subtype B env gene. J. Virol 89, 3380–3395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Punjani A, Rubinstein JL, Fleet DJ, and Brubaker MA (2017). cryo-SPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296. [DOI] [PubMed] [Google Scholar]
- Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, et al. ; MOPHTAVEG Investigators (2009). Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med 361, 2209–2220. [DOI] [PubMed] [Google Scholar]
- Sanders RW, Derking R, Cupo A, Julien J-P, Yasmeen A, de Val N, Kim HJ, Blattner C, de la Peña AT, Korzun J, et al. (2013). A next-gener ation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 9, e1003618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheres SHW (2012). RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol 180, 519–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simoncini D, Berenger F, Shrestha R, and Zhang KYJ (2012). A probabilistic fragment-based protein structure prediction algorithm. PLoS ONE 7, e38799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sok D, Pauthner M, Briney B, Lee JH, Saye-Francisco KL, Hsueh J, Ramos A, Le KM, Jones M, Jardine JG, et al. (2016). A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans. Immunity 45, 31–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suloway C, Pulokas J, Fellmann D, Cheng A, Guerra F, Quispe J, Stagg S, Potter CS, and Carragher B (2005). Automated molecular microscopy: the new Leginon system. J. Struct. Biol 151, 41–60. [DOI] [PubMed] [Google Scholar]
- Voss NR, Yoshioka CK, Radermacher M, Potter CS, and Carragher B (2009). DoG Picker and TiltPicker: software tools to facilitate particle selection in single particle electron microscopy. J. Struct. Biol 166, 205–213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- West AP Jr., Scharf L, Scheid JF, Klein F, Bjorkman PJ, and Nussenzweig MC (2014). Structural insights on the role of antibodies in HIV-1 vaccine and therapy. Cell 156, 633–648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu L, and Guan Y (2014). Immunologic Basis for Long HCDR3s in Broadly Neutralizing Antibodies Against HIV-1. Front. Immunol 5, 250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan M, Cottrell CA, Ozorowski G, van Gils MJ, Kumar S, Wu NC, Sarkar A, Torres JL, de Val N, Copps J, et al. (2019). Conformational Plasticity in the HIV-1 Fusion Peptide Facilitates Recognition by Broadly Neutralizing Antibodies. Cell Host Microbe 25, 873–883.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng SQ, Palovcak E, Armache J-P, Verba KA, Cheng Y, and Agard DA (2017). MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou T, Zheng A, Baxa U, Chuang GY, Georgiev IS, Kong R, O’Dell S, Shahzad-Ul-Hussan S, Shen CH, Tsybovsky Y, et al. ; NISC Comparative Sequencing Program (2018). A Neutralizing Antibody Recognizing Primarily N-Linked Glycan Targets the Silent Face of the HIV Envelope. Immunity 48, 500–513.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
3D EM reconstructions have been deposited in the Electron Microscopy Databank (http://www.emdataresource.org/) or Protein Databank (http://www.rcsb.org) under the accession numbers listed in the Key Resources Table.
KEY RESOURCES TABLE
| REAGENT OR RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Biological Samples | ||
| Plasma or serum from rhesus macaques | (Pauthner et al., 2017, Pauthner et al., 2018) | N/A |
| Chemicals, Peptides, and Recombinant Proteins | ||
| BG505 SOSIP.664 | Produced in house (Sanders et al., 2013) | N/A |
| 398F1 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| 25710 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| CE1176 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| X1632 SOSIP.664 | Produced in house (deCamp et al., 2014) | N/A |
| Protein A Resin | GE Healthcare | Cat# 17-5280-02 |
| Superose 6 Increase Size Exclusion Column | GE Healthcare | Cat# 17517201 |
| Uranyl Formate | Electron Microscopy Sciences | Cat# D310 25 GM |
| Lauryl Maltose Neopentyl Glycol | Anatrace | Cat# NG310 5 GM |
| Critical Commercial Assays | ||
| Pierce Fab Preparation Kit | Thermo Fisher Scientific | Cat# 44985 |
| Negative stain EM Grids | Electron Microscopy Sciences | Cat# EMS400-CU |
| Cryo-EM grids | Electron Microscopy Sciences | Cat# Q4100CR2 |
| Deposited Data | ||
| Cryo-EM Reconstruction of Polyclonal Serum Fab from a Protected Rhesus Macaque 4O9 Complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20396 |
| Negative-stain EM map of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20399 |
| Negative-stain EM map of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 | EMDataBank | EMD: 20400 |
| Negative-stain EM map of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20401 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–060 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20402 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12–065 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20403 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque OQ7 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20404 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12–153 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20405 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12M-248 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20409 |
| Negative-stain EM map of BG505-immunized and SHIV-challenged rhesus macaque 12–149 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20410 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–143 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20415 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20423 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12M-169 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20424 |
| Negative-stain EM map of BG505-immunized rhesus macaque 11M-088 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20425 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20426 |
| Negative-stain EM map of BG505-immunized rhesus macaque 4O9 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20427 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20428 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20946 |
| Negative-stain EM map of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20429 |
| Negative-stain EM map of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20970 |
| Negative-stain EM map of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20971 |
| Negative-stain EM map of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 | EMDataBank | EMD: 20972 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–084 wk 26 serum fab complexed with 398F1 SOSIP.664 | EMDataBank | EMD: 20992 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–137 wk 26 serum fab complexed with 398F1 SOSIP.664 | EMDataBank | EMD: 20936 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–046 wk 26 serum fab complexed with 398F1 SOSIP.664 | EMDataBank | EMD: 20937 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20973 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20974 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ05 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20975 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20976 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20977 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ11 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20978 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20979 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20980 |
| Negative-stain EM map from 3D sorting of unimmunized control rhesus macaque BZ13 (challenged with SHIV) serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20981 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–060 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20982 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–060 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20983 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–065 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20938 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–065 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20984 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–065 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20985 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque OQ7 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20939 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque OQ7 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20940 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–143 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20941 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–143 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20987 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20988 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20989 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20942 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20921 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–046 wk 0 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20943 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20944 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20899 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20898 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20897 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20896 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 12–137 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20896 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20895 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20894 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 12 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20893 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20990 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20892 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20891 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 11M-088 wk 28 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20890 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20889 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20889 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 10 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20888 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20887 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20991 |
| Negative-stain EM map from 3D sorting of BG505-immunized rhesus macaque 4O9 wk 26 serum fab complexed with BG505 SOSIP.664 trimer | EMDataBank | EMD: 20886 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–046 wk 26 serum fab complexed with 25710 | EMDataBank | EMD: 20885 |
| Negative-stain EM map of BG505-immunized rhesus macaque 12–084 wk 28 serum fab complexed with BG505 SOSIP trimer | EMDataBank | EMD: 20884 |
| PDB Coordinates of BG505 SOSIP.664 trimer complexed with Polyclonal Serum Fab from a Protected Rhesus Macaque 4O9 | Protein Data Bank | PDB ID: 6V0R |
| Experimental Models: Cell Lines | ||
| Human: FreeStyle HEK293F | Thermo Fisher Scientific | Cat# R79007 |
| Human: HEK294T | ATCC | Cat# CRL-3216 |
| Software and Algorithms | ||
| Unicorn 7.0 | GE Healthcare | https://www.gelifesciences.com/ |
| UCSF Chimera | Pettersen et al., 2004 | N/A |
| Appion database | Lander et al., 2009 | N/A |
| Leginon | Suloway et al., 2005 | N/A |
| DoG Picker | Voss et al., 2009 | N/A |
| Relion | Scheres, 2012 | N/A |
| CryoSparc | Punjani et al., 2017 | N/A |