

Abstract
Patients with damage to the orbitofrontal cortex display a variety of impairments in cognitive and affective function, including a reduced ability to use information about the consequences of their actions to guide their behavior. In this study, rats with neurotoxic lesions of the orbitofrontal cortex (OFC) failed to use specific expectancies about outcomes to guide their learning of an instrumental discrimination task. By contrast, lesioned rats were unimpaired in a measure of learned motivational function, the potentiation of feeding under conditions of food satiation, by a conditioned stimulus (CS) that had been paired with food while the rats were food-deprived. Notably, performance of both of these tasks has been shown to depend on function of the basolateral amygdala (BLA), a region that is richly interconnected with OFC. Thus, the present results are consistent with the view that the acquisition and use of specific outcome expectancies to guide behavior critically involves a neural system that includes ABL and OFC, but indicate that certain motivational properties acquired by cues on the basis of appetitive learning involve ABL circuitry apart from OFC.
Keywords: orbitofrontal cortex, outcome expectancy, potentiated feeding, associative learning, motivation, basolateral amygdala
Associative learning has consequences beyond the conditioning of overt motor behavior. Pairing an initially neutral conditioned stimulus (CS) with a reinforcer allows that CS to activate a memorial representation of that reinforcer (“outcome expectancy”), which can have many behavioral effects (Rescorla, 1988). Likewise, associative learning endows CSs with motivational properties. For example, CSs paired with food delivery often acquire the ability to serve as conditioned reinforcers for the establishment of new learning (Rescorla, 1980), and to modulate the performance of consummatory (Weingarten, 1983) or instrumental (Dickinson et al., 2000) behavior.
Recent studies in rats and monkeys suggest that the orbitofrontal cortex (OFC) is critical to using learned outcome expectancies to guide behavior (Izquierdo et al., 2004; Pickens et al., 2003), and to some aspects of learned motivational function (Pears et al., 2003). A common assessment of expectancy is the reinforcer devaluation procedure. In one version of this task, a rat is given light-food pairings, until the light consistently controls behavior directed to a food cup. Next, an aversion is established to the food by pairing it with an illness-inducing agent in the absence of the light. Finally, responding to the light is tested in the absence of the food. Whereas normal rats show a spontaneous reduction in food-cup behavior, rats with OFC lesions fail to show such a reduction (Gallagher et al., 1999; Pickens et al., 2003, 2005). This failure is thought to reflect an inability of OFC-lesioned animals to use a CS-activated expectancy of the newly-devalued reinforcer to guide responding.
At the same time, Pears et al. (2003) reported that OFC lesions interfered with learned motivational processes in marmosets. Although lesioned animals showed normal acquisition of instrumental responding reinforced by food, they showed significant deficits in acquiring responding that was reinforced by a previously-trained CS. The failure of a CS to serve as a conditioned reinforcer for instrumental responding in OFC-lesioned animals suggested that the OFC plays a critical role in the assignment of motivational value to the CS.
Similar impairments in conditioned reinforcement and in tests for the use of outcome expectancies, such as reinforcer devaluation, are also seen after lesions of the basolateral amygdala (ABL) (Burns et al., 1993; Hatfield et al., 1996). Because these two regions share rich reciprocal connections (Ghashghaie & Barbas, 2002), OFC and ABL form a circuit that may mediate both learned motivational functions and the use of outcome expectancies to guide behavior.
The experiments reported here sought to extend such findings by evaluating the effects of OFC lesions on the performance of rats in two additional associative learning tasks, CS-potentiated feeding (Weingarten, 1983), thought to engage learned motivational processes (Experiment 1), and a differential-outcome expectancy (DOE) task (Trapold & Overmier, 1972) in which instrumental performance is mediated by learned expectancies (Experiment 2). These tasks were selected on the basis of previous research showing them to be sensitive to disruptions in ABL function (Blundell et al., 2001; Holland et al., 2002).
Methods
Subjects
The subjects were 26 male Long-Evans rats (Charles River Laboratories, Raleigh, NC), which weighed 300–325 g when they arrived in the laboratory vivarium. After one week with ad lib access to food and water in individual cages, the rats were reduced to 85% of their ad libitum weights by restricting their access to food. The rats lived in individual cages, with free access to water, in a colony room illuminated from 6 a.m. to 8 p.m. Experiment 1 was conducted in two replications, one with 18 rats and one with 8 rats, with lesion and behavioral training procedures balanced across replications.
Surgical procedures
Aseptic surgeries to make bilateral neurotoxic lesions of OFC were performed under isoflurane (Isovet; Mallinckrodt, Mundelein, IL) anesthesia. The lesions were made using NMDA at a concentration of 20 mg/ml in PBS (Sigma, St Louis, MO), injected using a glass micropipette attached by a length of plastic tubing to a picospritzer (General Valve Corporation, Fairfield, NJ). OFC lesions were made in 16 rats, using 4 injection sites in each hemisphere: two injections at 4.0 mm anterior to bregma, 4.2 mm ventral from the skull surface at bregma, and placed 2.2 mm (0.08 μl ) and 3.7 mm (0.08 μl) from the midline, and 2 injections at 3.0 mm anterior to bregma, 5.2 mm ventral from the skull surface at bregma, and placed 4.2 mm (0.08 μl) and at 3.2 mm (0.05 μl) from midline. For 10 control rats, the glass micropipette was lowered to the same sites, but no injections were given.
Apparatus
The behavioral training apparatus for Experiment 1 consisted of four individual chambers (22.9 × 20.3 × 20.3 cm) with aluminum front and back walls, clear acrylic sides and top, and a floor made of 0.48-cm stainless steel rods spaced 1.9 cm apart. A dimly illuminated food cup was recessed in the center of one end wall. An infrared photocell placed just inside the food cup was polled (1 kHz) by computer circuitry. Each chamber was enclosed in a sound-resistant shell. A speaker, used to present the auditory CSs, was mounted on the inside wall of the inner shell, 10 cm above the experimental chamber and even with the end wall opposite the food cup. Ventilation fans provided masking noise (70 dB). Constant dim illumination was provided by a 6-w lamp behind a dense red lens, mounted next to the speaker. A TV camera was mounted within each shell to provide a view of the chamber. Television images were recorded in selected sessions, but the results of analyzing these recordings are not presented. The apparatus used in Experiment 2 comprised 8 chambers similar to those used in Experiment 1. These chambers included 3 visual CS sources, a jeweled 6-w lamp 9.5 cm over the food cup, and two 6-w lamps located on opposite side walls of the sound-attenuating shell, each centered 10 cm above the experimental chamber. In addition, any of three operant response devices could be inserted into each chamber. A 2×2 cm lever could be positioned 5 cm above the floor, 2.5 cm to the left of the food cup, a 10-cm chain could be suspended from the ceiling to within 10 cm of the floor, 3.5 cm to the left of the food cup, and rats could be given access to a 2.5 cm port with a paddle behind it, 2.5 cm to the right and 9 cm above the food cup.
Behavioral training procedures
Experiment 1 examined the performance of sham-lesioned and OFC-lesioned rats in a potentiated feeding procedure, which assessed the ability of a CS to promote feeding while the rats were food-sated. Hungry rats were first trained with a Pavlovian discrimination procedure, in which one auditory CS was paired with food and another was nonreinforced. After an extended period of food satiation, food consumption in the presence of the previously reinforced and nonreinforced CSs was examined. In previous studies, normal rats ate more during the reinforced CS than during the nonreinforced CS or during baseline periods (e.g., Holland et al., 2002; Holland & Gallagher, 2003; Petrovich et al., 2002), but rats with lesions of ABL did not show this potentiation of feeding. Those studies also showed that the enhanced consumption during reinforced CSs was independent of food-cup approach responses conditioned to the CSs: feeding was potentiated regardless of where food was placed in the chamber, and brain lesions that eliminated potentiated feeding had no effect on approach to the food cup, even in tests in which food was present in that cup.
All training sessions were 32 min duration and were given daily at approximately the same time (9 a.m.). The rats first received two sessions to train them to eat from the recessed food cup. In each of these sessions there were 8 deliveries of the reinforcer used throughout Experiment 1, 2 45-mg food pellets (Precision Diets, formula A), 0.5-s apart. In these and all remaining training sessions, the intertrial intervals were variable (mean = 4 min) within a range of 2 to 6 min. The rats then received training sessions to establish a discrimination between two auditory CSs. In each session of the first part of this training, half of the rats received 8 reinforced 10-s presentations of an 80-db white noise and the other half received 8 reinforced presentations of a 10-s, 80-db 1,500-hz tone. In the second portion of this training, the rats received 2 reinforced presentations of the CS they had received in the first part of training (CS+), randomly intermixed with 6 nonreinforced presentations of the other auditory stimulus (CS−). Due to experimenter error, the rats in the first replication received 2 initial CS+ training sessions and 12 discrimination training sessions, and the rats in the second replication received 4 initial CS+ training sessions and 8 discrimination training sessions.
The rats were then given 9 days free access to food in their home cages. On each of the last two of those days, the rats received a consumption test in the experimental chambers. Each consumption test began with a 10-min pretest of pellet consumption in the chambers before the presentation of CSs. In previous studies of potentiated feeding, we found that when satiated rats were placed in the experimental chambers, they often ran immediately to the food cup and ate any food there. The pretest was intended to reduce the contribution of this effect to feeding during the CS presentations. In the pretest, 50 pellets were present in the recessed food cup when the rats were placed in the chambers. After 10 min had elapsed, the rats were removed from the chambers and placed in transport cages. Food pellets were suctioned from the food cup and trays beneath the chamber floor and saved for counting. The food cups were then quickly refilled with 50 new pellets, and the rats were returned to the chambers for a 10 min test of consumption in the presence of CS+ or CS−. During that test, ten 10-s CS+s or CS−s were presented (but no additional pellets were delivered after CS+). Both the reinforcement contingency in training (CS+ or CS−) and identity of the CS (tone or noise) were counterbalanced over the two test sessions. Finally, at the end of the 10-min CS consumption test, the rats were quickly removed, and the remaining pellets suctioned and reserved for counting. Behavior was video recorded throughout each test, but analyses of behavior are not presented here.
Experiment 2 examined the performance of these same rats in an instrumental discrimination task in which specific outcome expectancies controlled by discriminative stimuli could be used to guide instrumental behavior. Previous studies showed that instrumental discriminations that involve multiple stimuli, responses, and reinforcers (outcomes) are learned more rapidly if each stimulus-response combination is consistently related to a unique outcome (Blundell et al., 2001; Trapold & Overmier, 1972), than if the stimulus-response combinations are randomly reinforced with both outcomes. This observation has been attributed to subjects’ use of learned outcome expectancies to supplement the explicit stimuli as cues to guide behavior. Consider a discrimination in which one response (R1) is reinforced only in the presence of one stimulus (S1), and another response (R2) is reinforced only in the presence of a second stimulus (S2). If correct responding during each stimulus is, in addition, associated with a unique outcome, then the unique outcome expectancies that each stimulus elicits can serve as supplemental discriminative cues to signal the correct response: only one response is reinforced in the presence of each expectancy. By contrast, if correct responding during each stimulus is randomly associated with both outcomes, then each response is reinforced in the presence of both expectancies, detracting from the ability of the explicit cues (S1 and S2) to control correct responding via differential outcome expectancies.
In Experiment 2, each rat was given training that involved two different reinforcement outcomes (flavored sucrose solutions), 3 different visual CSs, and 3 different operant responses. As indicated in Table 1, two of the stimulus-response combinations consistently led to unique outcomes (S1-R1-O1 and S2-R2-O2), whereas the third stimulus-response combination was reinforced with each outcome 50% of the time (S3-R3-O1/O2).
Table 1.
Differential Outcome Expectancy Test Procedures of Experiment 2
| Stimulus | Correct Response→Outcome | Errors |
|---|---|---|
| S1 | R1→O1 | R2-, R3- |
| S2 | R2→O2 | R1-, R3- |
| S3 | R3→O1/O2 | R1-, R2- |
Note. In each of the 5 sessions of discrimination training, 3 discriminative stimuli were presented and 3 response devices were available. O1 and O2 were two flavored sucrose solutions used as reinforcing outcomes (counterbalanced). R1 was a paddle-push response, R2 and R3 were lever-press and chain-pull responses (counterbalanced); bold-face type indicates the different-outcome error (see text). S1 was the illumination of a visual stimulus on one side of the chamber, and S2 and S3 were two other visual stimuli (counterbalanced). → indicates a response-outcome contingency. - indicates no scheduled consequence of a response.
The identities of O1 and O2 (0.3 ml of a solution of 1g of either orange or grape unsweetened Kool-Aid, and 50g sucrose in 1 l water), S2 and S3 (illumination of the jeweled lamp, flashing at a rate of 3 Hz, or the steady illumination of the lamp on the left side of the chamber), and R2 and R3 (lever press or chain pull) were completely counterbalanced. S1 was the illumination of the lamp on the right side of the chamber and R1 was paddle-pushing for all rats. Performance of an inappropriate response (e.g., R1 anytime other than during S1) had no consequence.
Rats were first shaped to perform each of the 3 operant responses needed for differential outcome expectancy training. In each shaping session only one of the operant responses was available. To encourage contact with the response device in these initial training sessions, it was covered with a paste of sucrose, chow dust and water. In the first session, performance of each R2 was reinforced with O2. The session ended for each rat after it reached 50 responses. In the second session, each R3 was reinforced with O1, until 25 responses were emitted, and in the third session, each R3 was reinforced with O2 until 25 responses were emitted. In the fourth session, each R1 was reinforced with O1 until 50 responses were emitted.
Next, rats were trained to emit each of these responses only in the presence of a particular visual signal, over the course of 9 32-min sessions. In each of these sessions, only 1 response device was available. In the first three sessions, R1 was reinforced (with O1) during 16 30-s presentations of S1; in the next 3 sessions, R2 was reinforced (with O2) during 16 30-s presentations of S2; and in the final 3 of these sessions, R3 was reinforced (50% with O1 and 50% with O2) during the 30-s presentation of S3. Any rats that failed to perform a response were given an extra operant response training session without signals on that response before the next training session.
In the DOE test phase (shown in Table 1), the rats received 5 30-min sessions of discrimination training, in which all three responses devices were available, and all three 30-s signals were presented in each session. In each of these sessions, the rats received 8 S1-R1-O1 trials, in which each R1 during S1 was reinforced with O1, 8 S2-R2-O2 trials, in which each R2 during S2 was reinforced with O2, and 8 S3-R3-O1/O2 trials, in which each R3 during S3 was reinforced with either O1 (4 trials) or O2 (4 trials). The performance of any response other than the correct, reinforced response was recorded as an error, but had no consequence.
To provide evidence on the ability of the lesioned rats to discriminate the two reinforcers, at the conclusion of the experiment we administered a test of discriminative taste aversion learning to all of the rats. In each of two training days, separated by an empty day, each rat received 5 min access to 12 ml of either orange- or grape-flavored sucrose in the liquid cups, paired with a 5 ml/kg injection of 0.3M lithium chloride. Discriminative taste aversion learning was assessed by measuring the rats’ consumption of each of the two flavored sucrose solutions. On the second and third days after the final flavor-toxin pairing, the rats received 5-min consumption tests in which 12 ml of solution was made available in the liquid cups. The order of solution presentation was counterbalanced.
Response measures
Experiment 1: The measure of conditioning to the auditory CSs during the Pavlovian training phase was the percentage of time during which the food cup photobeam was broken (presumably indicating that a rat’s head was in the food cup) during the last 5 s of each CS interval. In addition, the percentage of time in the food cup during the 5-s intervals immediately before each trial was recorded as a measure of baseline responding. The measure of consumption was the number of food pellets eaten, which was measured by subtracting the number of whole pellets remaining in the food cups and trays from 50. In the rare case of a rat’s leaving fractional pellets uneaten, a fractional pellet was arbitrarily defined as 0.5 pellet. In no case did a rat leave more than two fractional pellets. We reported the number of pellets consumed in the pretest periods, and during the separate CS+ and CS− tests. The difference between the number of pellets consumed in CS+ tests and CS− tests served as a measure of the effect of the CS’s past conditioning relation with food on consumption in each rat, and thus served as a direct, within-subject, measure of the potentiated feeding effect.
Experiment 2: For each rat, we recorded the percentages of each trial type (S1, S2, or S3) on which each response (R1, R2, or R3) occurred, as well as the percentage of dummy trials (empty intervals equal in duration to the trials, immediately before each trial) on which each of those responses occurred. The primary measure of discriminative responding in the DOE test phase was responding during S1. The correct (reinforced) response during S1 was R1; the critical data were the likelihood of R2 and R3 errors during S1. If learned expectancies of O1 and O2 were used as discriminative cues to direct responding, then R3 errors would be more frequent than R2 errors, because R1 and R3 shared the outcome O1, whereas R1 and R2 produced different outcomes. Thus, for responding during S1, R2 was defined as the “different outcome error” (bold type in Table 1) and R3 was termed the “shared outcome error”. Comparable measures were constructed for responding during S2, in which R2 was the correct response, R1 was the different-outcome error (bold type in Table 1) and R3 was the shared-outcome error. However, we used responding during S1 as our primary measure, because S1 was the same stimulus for all rats, and S2, S3, R2, and R3 were perfectly counterbalanced among all rats.
Histological procedures
After completion of the behavioral testing in Experiment 2, the rats were deeply anaesthetized with Nembutal (150 mg/kg), and perfused with 0.1-M phosphate-buffered saline (PBS), followed by 10% (v/v) formalin. The brains were removed and stored in 0.1-M PBS with 20% (w/v) sucrose 4 C° for 24–48 hr. Sections (40-μm) were taken from each brain, and alternate sections were mounted on slides and Nissl-stained to verify the lesions.
Results
Histological results
Ten rats had acceptable lesions of OFC, encompassing the lateral and dorsolateral orbital regions, the lateral portions of ventral orbital cortex, and portions of both dorsal and ventral agranular insular cortex. This target region was designed to include areas on the dorsal bank of the rhinal sulcus that have reciprocal interactions with ABL (Kita & Kitae, 1990; Krettek & Price, 1977; Shi & Cassell, 1998), but to exclude medial orbital and the medial portions of ventral orbital cortex and gustatory regions located in agranular insular cortex posterior to the genu of the corpus callosum (Kosar et al., 1986a, 1986b; Krushel & Van Der Kooy, 1988; Saper, 1982). The lesions averaged 60% damage of the target region bilaterally. In most brains, damage was substantial along the anterior-posterior axis of the target region from 2.7 to 4.7 mm anterior to bregma. No lesion extended anterior to the coronal plane +5.0 or posterior to the plane +2.2 relative to bregma. Photomicrographs of a representative lesion and sham brain are shown in Figure 1. Six lesioned rats showed little evidence of damage and were discarded from the analyses. There was little discernable damage in any of the 10 sham-lesioned control rats.
Figure 1.

Photomicrographs showing typical sham (left hemisphere) and neurotoxic (right hemisphere) lesion of orbitofrontal cortex. The arrows outline the lesion.
Behavioral results
Experiment 1, Pavlovian discrimination training
Two lesioned rats and one sham rat failed to acquire the auditory discrimination; their data were discarded from Experiment 1, leaving 8 OFC-lesioned and 9 sham rats. The OFC lesion did not affect auditory Pavlovian discrimination learning, which was acquired rapidly in all remaining rats. The left panel of Figure 2 shows performance in the final four training sessions; a lesion (OFC or sham) X stimulus (reinforced or nonreinforced CS) ANOVA showed a significant effect of stimulus, F(1, 15) = 113.96, but no effect of lesion or lesion X stimulus interaction, Fs < 1.
Figure 2.

Left panel: Mean (±sem) food cup conditioned responding during the reinforced (CS+) and nonreinforced (CS−) conditioned stimuli over the last 4 sessions of Experiment 1. Right panel: Potentiated feeding consumption test results of Experiment 1. Each bar shows the mean difference (±sem) between pellet consumption during the test session that included the previously-reinforced CS and consumption during the test with the nonreinforced CS. OFC = orbitofrontal lesion.
Experiment 1: Consumption tests
The primary data of this experiment are the results of the consumption tests with CS+ and CS−. The right panel of Figure 2 shows consumption difference scores (CS+ minus CS−) for OFC-lesioned and sham-lesioned rats. Both sham and lesioned rats ate more pellets in the test in which the previously reinforced CS+ was presented (15.6 ± 5.9 and 19.4 ± 4.6 pellets, respectively) than in the test with the nonreinforced CS− (7.4 ± 4.3 and 11.6 ± 5.9 pellets). Thus, the ability of the noise and tone stimuli to potentiate eating depended on their associative history, that is, their prior pairing with food. A lesion X test (CS+ or CS−) ANOVA showed only a main effect of test (CS+ vs CS−), F(1, 15) = 15.85, p = .001; remaining Fs < 1. Likewise, a contrast of the difference scores portrayed in Figure 2B showed no effect of lesion, F < 1.
Pretest food consumption also did not differ as a function of the lesion, averaging 29.6 ± 4.2 pellets in OFC-lesioned rats and 30.4 ± 3.6 pellets in sham lesioned rats, F< 1. Finally, the lesions had no effect on the rats’ test weights or weight gain during the satiation phase, Fs < 1.
Experiment 2
Three OFC and 2 sham rats failed to acquire one or more of the 3 operants in the initial response shaping sessions; they were eliminated from Experiment 2, leaving 7 OFC and 8 sham rats. The lesions did not affect acquisition of responding under the control of the 3 visual signals during the initial conditioning sessions, in which only one signal and its correct response device was available. By the last of each of these sessions, the percentage of trials on which at least one response occurred was 84.4 ± 6.6%, 88.3 ±7.1%, and 90.6 ± 6.0% during S1, S2, and S3 respectively in the lesioned rats, and 86.6 ± 8.9%, 89.3 ±5.9%, and 88.4 ± 6.6% in the sham rats.
The primary data of Experiment 2 were the acquisition of the test discriminations under the conditions shown in Table 1. Figure 3 shows performance during S1, in which R1 responses were always reinforced with O1, and errors, consisting of R2 or R3 responses, were not reinforced. R2 responses were always reinforced with a different outcome (O2) on S2 trials, whereas R3 was reinforced with the same outcome (O1) as R1 on half of the S3 presentations, and with O2 on the other half of S3 trials. Over all 5 sessions (summarized in Figure 4a), sham rats showed more shared-outcome errors (R3) than different-outcome errors (R2; bold type in Table 1) during S1. This result can be attributed to the rats’ use of specific outcome expectancies to guide responding: responses reinforced in the presence of different expectancies (e.g., R1 and R2) were more readily differentiated than responses which yielded the same outcome (e.g., R1 and R3). By contrast, OFC-lesioned rats showed equivalent levels of shared-and different-outcome errors. The absence of any difference in shared- and different-outcome errors in these rats is consistent with an impairment in the associative basis for outcome expectancy learning.
Figure 3.
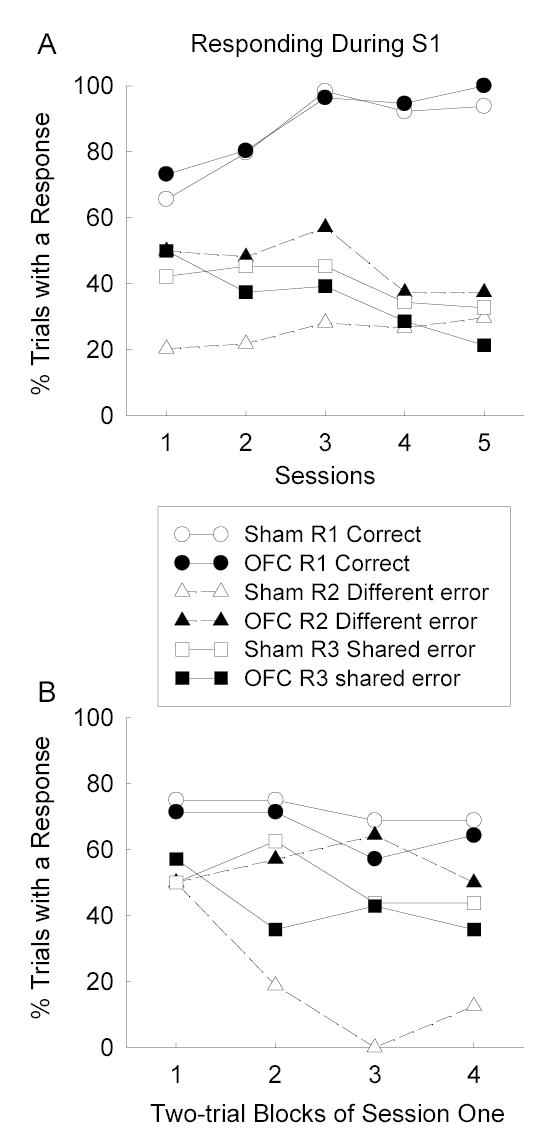
Acquisition of instrumental responding during stimulus S1 in the differential-outcome expectancy (DOE) test phase of Experiment 2. Panel A shows mean responding in each session and Panel B shows acquisition of responding within the first test session alone. The error bars provide measures of variance appropriate to the error terms for the lesion X error type interactions, 2(mean square error/df)1/2. OFC = orbitofrontal cortex.
Figure 4.

Overall mean (± sem) responding during stimulus S1 in the differential-outcome expectancy (DOE) test phase of Experiment 2. Panel A shows the percentage of trials on which each type of response occurred at any time during S1, and Panel B shows only responding that occurred prior to the delivery of the first reinforcer on each trial. OFC = orbitofrontal cortex.
The levels of correct R2 responses on S2 trials (86.6±10.9% in shams and 79.6±15.8% in lesioned rats) and R3 responses on S3 trials (88.1±10.0%, and 85±16.3%, respectively), did not differ among either responses or groups. Finally, Figure 3B, which portrays successive two-trial blocks of responding during the first session, shows that the differences in responding during the first training session reflected rapid acquisition of the differences in error frequencies, rather than different initial levels of those responses.
It might be argued that the sham rats were using cues provided by the reinforcing outcomes themselves to guide instrumental responding, rather than S1-evoked expectancies of those outcomes (e.g. Blundell et al., 2001). In that case, the deficit in the lesioned rats’ performance would reflect a deficit in their ability to use overt flavor cues, rather than flavor expectancies, to guide responding. To evaluate that possibility, we reanalyzed the data shown in Figure 4a, excluding responses that occurred after the delivery of the first reinforcer on each trial. The remaining responses could not have been cued by the physical reinforcer delivered on that trial, and thus more clearly reflect the influence of S1-evoked outcome expectancies. These data (Figure 4b) showed a very similar pattern to those from the entire stimulus duration (Figure 4a).
A lesion X error (R2 or R3) X sessions ANOVA of the data shown in Figure 3a (and summarized in Figure 4a) showed a significant lesion X error interaction, F(1, 13) = 6.47, p = .024. Individual post-hoc contrasts (Tukey HSD procedure, p < .05) showed that the sham rats showed significantly fewer R2 different-outcome errors than R3 shared-outcome errors, and also significantly fewer R2 different-outcome errors than the OFC-lesioned rats. No other contrast was significant. A comparable ANOVA of the data that excluded post-reinforcement responses (Figure 4b) also showed a reliable lesion X error interaction, F(1, 13) = 11.83, p = .004. As with the analysis of the overall data, individual Tukey HSD (ps < .05) contrasts showed that the sham rats showed significantly fewer R2 different-outcome errors than R3 shared-outcome errors, and also significantly fewer R2 different-outcome errors than the OFC-lesioned rats. No other contrast was significant, although the numerically lower levels of R3 errors in OFC-lesioned rats than in sham-lesioned rats approached significant, p = .082.
Comparable analyses of performance during S2 (not shown in Figures 3 or 4) were also performed. For those analyses, R2 was the correct response, R1 the different-outcome error and R3 the shared-outcome error. As with the analyses of S1 responding, sham-lesioned rats showed more shared-outcome (R3) errors (49±4.1%) than different-outcome (R1) errors (29.7±8.5%), but OFC-lesioned rats did not (39.3±6.9% and 39.9±8.1%). Statistical analyses like those performed on S1 responding yielded comparable outcomes.
Finally, although the lesioned rats showed no evidence of using differential outcome expectancies to guide their instrumental responding, they learned the final flavor aversion discrimination as well as shams. Thus, their deficiency in using outcome expectancies is not attributable to an inability to discriminate the two outcomes. The OFC-lesioned rats consumed means of 2.7 ± 0.5 ml of the devalued flavor and 6.8 ± 1.5 ml of the nondevalued flavor, and the sham-lesioned rats consumed 2.7 ± 0.7 and 7.2 ± 0.7 ml, respectively. A lesion X flavor ANOVA showed a significant effect of flavor (toxin-paired or not presented in training), F(1, 13) = 28.99, p < .001, but no effect of lesion or its interaction with flavor, Fs < 1.
Discussion
In this study, rats with neurotoxic or sham lesions of OFC were tested in both a CS-potentiated feeding task and a differential outcome expectancy (DOE) task. Although rats with OFC lesions showed normal CS-potentiated feeding in Experiment 1, their performance on the DOE task was substantially impaired in Experiment 2. In that experiment, rats were trained to make different cue-specific instrumental responses, which led to distinct reinforcing outcomes (two flavors of sucrose solution). Although the pattern of errors observed showed that sham-lesioned rats used cue-elicited outcome expectancies to help guide their instrumental responding, OFC-lesioned rats showed no evidence of using outcome expectancies to guide their behavior. This failure was not attributable to a deficit in discriminating the two outcomes themselves, because the lesioned rats showed no deficit in discriminating between the two flavored outcomes in a flavor aversion learning task conducted at the end of the study.
The OFC lesion deficit observed in Experiment 2 is consistent with many accounts of OFC function, including those that emphasize OFC’s role in the representation of information about rewarding outcomes (e.g., Montague & Burns, 2002; Rolls, 2000), those that focus on its role in response selection (e.g., Arana et al., 2003) and those that point to its involvement in response inhibition (Dias et al., 2000). Expectancy-guided behavior in the DOE task required learned associative access to specific representations of the two reinforcing outcomes by the visual cues, the selection of an instrumental response appropriate to the outcome anticipated on each trial, and the suppression of responses inappropriate to that outcome. Thus, in principle, OFC lesion-induced deficits in any of these processes might underlie the lesion effects observed in Experiment 2. However, it is notable that OFC-lesioned rats were no poorer than sham-lesioned rats at suppressing shared-outcome errors. Instead, lesioned rats were only deficient at using outcome expectancies to suppress the performance of a response (the different-outcome error) that was never appropriate to the anticipated outcome. Thus, the deficit observed here implicates OFC in the acquisition or use of specific outcome expectancies, rather than a more general inhibitory function.
OFC neurons have been demonstrated to selectively code properties of expected rewards, such as their magnitude and valence, in many experimental settings (e.g. Schoenbaum et al., 1998, 1999; Tremblay & Schultz, 1999). It is notable that the two outcomes used in Experiment 2 differed only in flavor; they had equal caloric value and were equally preferred. Thus, OFC was important for acquiring or using expectancies that coded sensory properties of the reinforcing outcomes. OFC is well-positioned for such a role, because it is richly interconnected with brain regions that process the gustatory and olfactory information necessary to distinguish the two outcomes used in Experiment 2, as well as visual information provided by the discriminative stimuli (e.g. Carmichael & Price, 1995a, b).
Holland and Gallagher (2004) noted that the acquisition of outcome expectancy information and the use of that information to guide behavior may depend not only on OFC function, but also on OFC’s interaction with ABL. For example, performance in the devaluation task (described in the introduction) is disrupted by lesions of either ABL (Balleine et al., 2003; Hatfield et al., 1996) or OFC (Gallagher et al., 1999; Pickens et al., 2003). Similarly, the impaired performance of OFC-lesioned rats in the DOE task of Experiment 2 complements observations of deficits in DOE performance after ABL lesions. Blundell et al. (2001) found that ABL lesions disrupted performance in a simpler, between-subjects version of the DOE task, and in an unpublished study, we found ABL lesion deficits using procedures identical to those of Experiment 2. Finally, Schoenbaum et al. (2003) found that a neural correlate of outcome expectancy that emerges in OFC in the presence of a predictive cue depends critically on ABL integrity. Such encoding failed to emerge in OFC recordings ipsilateral to a neurotoxic lesion in ABL when rats used different odor cues to predict the occurrence of either a positive (sucrose) or negative (quinine) reinforcer. Taken together, these data provide additional support for Holland and Gallagher’s (2004) view.
In contrast to the behavioral deficits produced by OFC lesions in Experiment 2, in Experiment 1 OFC damage had no effect on the ability of a Pavlovian CS, trained with a food reinforcer while the rats were food-deprived, to enhance feeding after a week of food satiation. Coupled with the results of Experiment 2, the failure to find OFC lesion effects in Experiment 1 is of interest in constraining the basis for CS-potentiated feeding. Specifically, associatively-activated representations of outcome information apparently do not play a critical role in CS-potentiated feeding. For example, it might otherwise be argued that a CS potentiates feeding while sated by activating a memorial representation of the food’s value when the rat was in the deprived state, at the time of original training. Because OFC damage interfered with associatively activated outcome representations in these same rats in Experiment 2, it is unlikely that such information was involved in the potentiated feeding observed with the procedures of Experiment 1.
Although we found no effects of OFC lesions on CS-potentiated feeding, others have suggested that OFC serves an important motivational function in associative learning, whereby previously neutral cues acquire motivational value as a result of pairings with reinforcers (e.g., Baxter & Murray, 2002; Pears et al., 2003). Furthermore, those authors suggested that this learned motivational function of OFC depends on its interaction with BLA, as we discussed earlier in the case of learning outcome expectancies. Consistent with this claim, although CSs paired with food typically acquire the ability to serve as conditioned reinforcers (Fantino, 1977; Rescorla, 1980), that ability is impaired in animals with lesions of either ABL (Burns et al., 1993; Cador et al., 1989; Hatfield et al., 1996; Parkinson et al., 2001) or OFC (Pears et al., 2003). It seemed reasonable to anticipate that another ABL-dependent motivational function acquired by CSs paired with food, the potentiation of feeding in satiation (Holland et al., 2002), would also critically involve OFC. However, the results of Experiment 1 indicate that ABL-OFC connectivity is not critical to this function.
This discrepancy between the effects of OFC lesions on potentiated feeding and on conditioned reinforcement has implications for our understanding of both ABL-OFC systems function and the nature of these two learning phenomena. Although both of these phenomena are described as reflecting acquired motivational functions of CSs, and both are dependent on ABL function, they differ in a number of important ways. For example, potentiated feeding involves enhancement of a species-typical consummatory response elicited by an innate reinforcer, whereas conditioned reinforcement involves assessment of a cue’s ability to support the acquisition or maintenance of an arbitrary instrumental response. It is possible that for a cue to serve as a conditioned reinforcer, it must maintain access to the value of the original primary reinforcer from which the conditioned reinforcement value was derived, a process known to be OFC-dependent (e,g., Pickens et al., 2003). At the same time, modulation of feeding itself by learned cues may involve more direct access to neural systems that control ingestive behavior. Thus the role of ABL in potentiated feeding may parallel somewhat the role of the amygdala in other learned control of innate behavioral systems, such as conditioned fear. Indeed, Petrovich et al. (2002) found that CS-potentiated feeding, but not conditioned reinforcement, depends on ABL projections to the lateral hypothalamus (LH). Notably, this pattern is opposite to that found for OFC (Exp. 1; Pears et al., 2003).
A better understanding of the basis for abnormalities often reported in patients with OFC damage (e.g., Bechara et al., 1997; Dolan, 1999) is emerging from the use of behavioral tasks as analytic tools in experimental animals. Such research is shedding light on how information processing within broader neural systems subserves functions that depend on OFC. The features of such OFC-dependent functions that can be modeled in a variety of tasks will help to provide a theoretical basis for understanding the role of this system in cognition and affect.
Acknowledgments
We thank Erin Kerfoot and Vanessa McKenna for technical support.
Footnotes
Supported by NIH grant MH53667. MAM and MPS contributed equally to this article.
References
- Arana FS, Parkinson JA, Hinton E, Holland AJ, Owen AM, Roberts AC. Dissociable contribution of the human amygdala and orbitofrontal cortex to incentice motivation and goal selection. J Neurosci. 2003;23:9632–9638. doi: 10.1523/JNEUROSCI.23-29-09632.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balleine BW, Killcross AS, Dickinson A. The effect of lesions of the basolateral amygdala on instrumental conditioning. J Neurosci. 2003;23:666–675. doi: 10.1523/JNEUROSCI.23-02-00666.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baxter MG, Murray EA. The amygdala and reward. Nature Rev: Neurosci. 2002;3:563–573. doi: 10.1038/nrn875. [DOI] [PubMed] [Google Scholar]
- Bechara A, Darmasio H, Tranel D, Damasio AR. Deciding advantageously before knowing the advantageous strategy. Science. 1997;275:1293–1294. doi: 10.1126/science.275.5304.1293. [DOI] [PubMed] [Google Scholar]
- Blundell P, Hall G, Killcross S. Lesions of the basolateral amygdala disrupt selective aspects of reinforcer representation in rats. J Neurosci. 2001;21:9018–9026. doi: 10.1523/JNEUROSCI.21-22-09018.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns LH, Robbins TW, Everitt BJ. Differential effects of excitotoxic lesions of the basolateral amygdala, ventral subiculum and medial prefrontal cortex on responding with conditioned reinforcement and locomotor activity potentiated by intra-accumbens infusions of D-amphetamine. Behav Brain Res. 1993;55:167–183. doi: 10.1016/0166-4328(93)90113-5. [DOI] [PubMed] [Google Scholar]
- Cador M, Robbins TW, Everitt BJ. Involvement of the amygdala in stimulus-reward associations: interaction with the ventral striatum. Neuroscience. 1989;30:77–86. doi: 10.1016/0306-4522(89)90354-0. [DOI] [PubMed] [Google Scholar]
- Carmichael ST, Price JL. Limbic connections of the orbital and medial prefrontal cortex in macaque monkeys. J Comp Neurol. 1995a;363:615–641. doi: 10.1002/cne.903630408. [DOI] [PubMed] [Google Scholar]
- Carmichael ST, Price JL. Sensory and premotor connections of the orbital and medial prefrontal cortex of macaque monkeys. J Comp Neurol. 1995b;363:642–664. doi: 10.1002/cne.903630409. [DOI] [PubMed] [Google Scholar]
- Dias R, Robbins TW, Roberts AC. Dissociable forms of inhibitory control within prefrontal cortex with an analog of the Wisconsin Card Sort Test: restriction to novel situations and independence from “on-line” processing. J Neurosci. 2000;17:9285–9297. doi: 10.1523/JNEUROSCI.17-23-09285.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickinson A, Smith J, Mirenowicz J. Dissociation of Pavlovian and instrumental incentive learning under dopamine antagonists. Behav Neurosci. 2000;114:468–483. doi: 10.1037//0735-7044.114.3.468. [DOI] [PubMed] [Google Scholar]
- Dolan RJ. On the neurology of morals. Nature Neurosci. 1999;2:927–929. doi: 10.1038/14707. [DOI] [PubMed] [Google Scholar]
- Fantino E (1977) Conditioned reinforcement: Choice and information. In WK Honig & JER Staddon (Eds) Handbook of operant behavior (313–339). Englewood Cliffs, NJ: Prentice-Hall.
- Gallagher M, McMahan RW, Schoenbaum G. Orbitofrontal cortex and representation of incentive value in associative learning. J Neurosci. 1999;19:6610–6614. doi: 10.1523/JNEUROSCI.19-15-06610.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghashghaei HT, Barbas H. Pathways for emotion: interactions of prefrontal and anterior temporal pathways in the amygdala of the rhesus monkey. Neurosci. 2002;115:1261–1279. doi: 10.1016/s0306-4522(02)00446-3. [DOI] [PubMed] [Google Scholar]
- Hatfield T, Han JS, Conley M, Gallagher M, Holland P. Neurotoxic lesions of basolateral, but not central, amygdala interfere with Pavlovian second-order conditioning and reinforcer devaluation effects. J Neurosci. 1996;16:5256–5265. doi: 10.1523/JNEUROSCI.16-16-05256.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holland PC, Gallagher M. Double dissociation of the effects of lesions of basolateral and central amygdala on CS-potentiated feeding and Pavlovian-instrumental transfer. Eur J Neurosci. 2003;17:1680–1694. doi: 10.1046/j.1460-9568.2003.02585.x. [DOI] [PubMed] [Google Scholar]
- Holland PC, Gallagher M. Amygdala-frontal interactions and reward expectancy. Curr Opin Neurobio. 2004;14:148–155. doi: 10.1016/j.conb.2004.03.007. [DOI] [PubMed] [Google Scholar]
- Holland PC, Petrovich GD, Gallagher M. The effects of amygdala lesions on conditioned stimulus-potentiated eating in rats. Physiol Behav. 2002;76:117–129. doi: 10.1016/s0031-9384(02)00688-1. [DOI] [PubMed] [Google Scholar]
- Izquierdo A, Suda RK, Murray EA. Bilateral orbital prefrontal cortex lesions in rhesus monkeys disrupt choices guided by both reward value and reward contingency. J Neurosci. 2004;24:7540–7548. doi: 10.1523/JNEUROSCI.1921-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kita H, Kitae ST. Amygdaloid projections to the frontal cortex and the striatum in the rat. J Comp Neurol. 1990;298:40–49. doi: 10.1002/cne.902980104. [DOI] [PubMed] [Google Scholar]
- Kosar E, Grill HJ, Norgren R. Gustatory cortex in the rat. I. Physiological properties and cytoarchitecture. Brain Res. 1986a;379:329–341. doi: 10.1016/0006-8993(86)90787-0. [DOI] [PubMed] [Google Scholar]
- Kosar E, Grill HJ, Norgren R. Gustatory cortex in the rat. II. Thalmocortical projections. Brain Res. 1986b;379:342–352. doi: 10.1016/0006-8993(86)90788-2. [DOI] [PubMed] [Google Scholar]
- Krettek JE, Price JL. Projections from the amygdaloid complex to the cerebral cortex and thalamus in the rat and cat. J Comp Neurol. 1977;172:687–722. doi: 10.1002/cne.901720408. [DOI] [PubMed] [Google Scholar]
- Krushel LA, Van der Kooy D. Visceral cortex: integration of the mucosal sense with limbic information in the rat agrangular insular cortex. J Comp Neurol. 1988;270:39–54. doi: 10.1002/cne.902700105. [DOI] [PubMed] [Google Scholar]
- Montague PR, Berns GS. Neural economics and the biological substrates of valuation. Neuron. 2002;36:265–284. doi: 10.1016/s0896-6273(02)00974-1. [DOI] [PubMed] [Google Scholar]
- Parkinson JA, Crofts HS, McGuigan M, Tomic DL, Everitt BJ, Roberts AC. The role of the primate amygdala in conditioned reinforcement. J Neurosci. 2001;21:7770–7780. doi: 10.1523/JNEUROSCI.21-19-07770.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pears A, Parkinson JA, Hopewell L, Everitt BJ, Roberts AC. Lesions of the orbitofrontal but not medial prefrontal cortex disrupt conditioned reinforcement in primates. J Neurosci. 2003;23:11189–11201. doi: 10.1523/JNEUROSCI.23-35-11189.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrovich GD, Setlow B, Holland PC, Gallagher M. Amygdalo-hypothalamic circuit allows learned cues to override satiety and promote eating. J Neurosci. 2002;22:8748–8753. doi: 10.1523/JNEUROSCI.22-19-08748.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickens CL, Saddoris MP, Setlow B, Gallagher M, Holland PC, Schoenbaum G. Different roles for orbitofrontal cortex and basolateral amygdala in a reinforcer devaluation task. J Neurosci. 2003;23:11078–11084. doi: 10.1523/JNEUROSCI.23-35-11078.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickens CL, Saddoris MP, Gallagher M, Holland PC. Orbitofrontal lesions impair use of cue-outcome associations in a devaluation task. Behav Neurosci. 2005;119:317–322. doi: 10.1037/0735-7044.119.1.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rescorla RA (1980) Pavlovian Second order conditioning: Studies in associative learning. Hillsdale, NJ: Erlbaum.
- Rescorla RA. Pavlovian conditioning : It’s not what you think it is. Amer Psychol. 1988;43:151–160. doi: 10.1037//0003-066x.43.3.151. [DOI] [PubMed] [Google Scholar]
- Rolls ET. The orbitofrontal cortex and reward. Cerebral Cortex. 2000;10:284–294. doi: 10.1093/cercor/10.3.284. [DOI] [PubMed] [Google Scholar]
- Saper CB. Convergence of autonomic and limbic connections in the insular cortex of the rat. J Comp Neurol. 1982;210:163–173. doi: 10.1002/cne.902100207. [DOI] [PubMed] [Google Scholar]
- Schoenbaum G, Chiba AA, Gallagher M. Orbitofrontral cortex and basolateral amygdala encode expected outcomes during learning. Nature Neurosci. 1998;1:155–159. doi: 10.1038/407. [DOI] [PubMed] [Google Scholar]
- Schoenbaum G, Chiba AA, Gallagher M. Neural encoding in orbitofrontal cortex and basolateral amygdala during olfactory discrimination learning. J Neurosci. 1999;19:1876–1884. doi: 10.1523/JNEUROSCI.19-05-01876.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoenbaum G, Setlow B, Saddoris MP, Gallagher M. Encoding predicted outcome and acquired value in orbitofrontal cortex during cue sampling depends upon input from basolateral amygdala. Neuron. 2003;39:855–867. doi: 10.1016/s0896-6273(03)00474-4. [DOI] [PubMed] [Google Scholar]
- Shi CJ, Cassell MD. Cortical, thalamic, and amygdaloid connections of the anterior and posterior insular cortices. J Comp Neurol. 1998;399:440–468. doi: 10.1002/(sici)1096-9861(19981005)399:4<440::aid-cne2>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- Trapold MA, Overmier JB (1972) The second learning process in instrumental training. In A Black & W F Prokasy (Eds), Classical Conditioning II, 427–452. New York: Appleton-Century-Crofts.
- Tremblay L, Schultz W. Relative reward preference in primate orbitofrontal cortex. Nature. 1999;398:704–708. doi: 10.1038/19525. [DOI] [PubMed] [Google Scholar]
- Weingarten HP. Conditioned cues elicit feeding in sated rats: A role for learning in meal initiation. Science. 1983;220:431–433. doi: 10.1126/science.6836286. [DOI] [PubMed] [Google Scholar]